
Археи Asgard — счастливые обладатели актинового цитоскелета
10 февраля 2023
Археи Asgard — счастливые обладатели актинового цитоскелета
- 1032
- 1
- 4
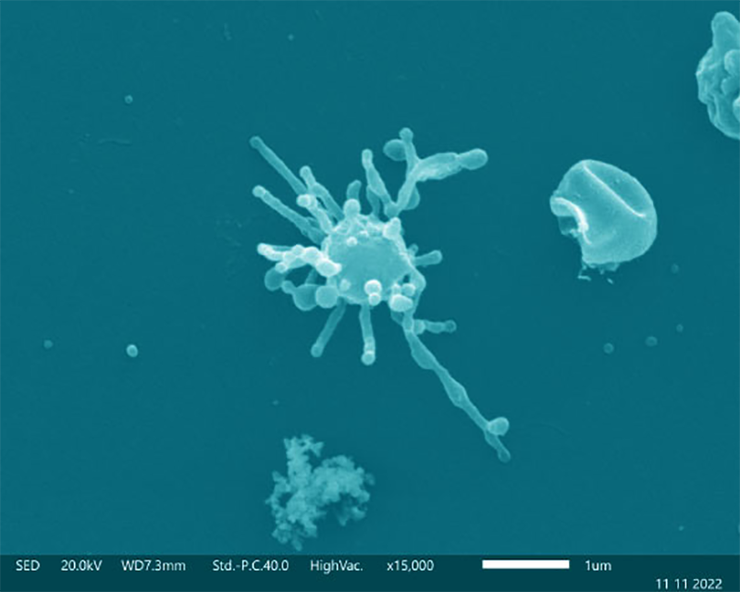
Ученым удалось вырастить в культуре еще одну архею из группы Asgard, Candidatus Lokiarchaeum ossiferum. Изображение получено с помощью сканирующей электронной микроскопии.
Рисунок в полном размере.
-
Автор
-
Редакторы
Загадочные археи Asgard, которых принято считать предками эукариот, с трудом поддаются культивированию. О существовании подавляющего большинства из них известно лишь по геномным последовательностям, полученным с помощью метагеномики, и до последнего времени в лабораторной культуре удалось вырастить лишь один вид. Недавно в Nature появилось сообщение о получении культуры еще одной асгардархеи, причем ее всестороннее изучение позволило узнать немало нового о биологии наших далеких пращуров...
После того, как в 1980-х годах археи были выделены Карлом Везе в отдельный домен, наряду с бактериями и эукариотами, на основании последовательностей 16S рРНК, стало понятно, что археи и эукариоты, скорее всего, имеют общее происхождение (рис. 1).

Рисунок 1. Древо жизни, построенное по последовательностям 16S рРНК. Можно видеть, что домены архей и эукариот кластеризуются в одну группу, сестринскую по отношению к бактериям.
Окончательно внести ясность в этот вопрос удалось существенно позже, когда в руках ученых благодаря метагеномике оказались геномы микроорганизмов, которые никто никогда не получал в виде лабораторной культуры. Среди них были и необычные археи, обладающие рядом свойств, присущих только эукариотам. В 2015 году в журнале Nature появилось сообщение о выделении группы архей Lokiarchaeota (сейчас она известна как Lokiarchaeia), которая формирует монофилетическую кладу с эукариотами [1]. В геномах ее представителей закодирован ряд белков, считавшихся уникальными для эукариот. Похожие белки участвуют в перестройках мембран, их еще называют сигнатурными белками эукариот. Это открытие, казалось бы, должно было поставить точку в спорах относительно общности происхождения и эволюционных взаимоотношений архей и эукариот. Некоторые ученые, однако, восприняли результаты исследования с долей скептицизма. В самом деле, этих таинственных локиархей никто не видел, а геномы, собранные из фрагментов ДНК в морской воде с помощью метагеномики, можно счесть артефактами работы биоинформатических программ. На протяжении нескольких лет аргументированно возразить таким скептикам было невозможно: ни одна лаборатория в мире на тот момент не сообщила о том, что ей удалось вырастить локиархей в культуре. Впрочем, это обстоятельство не мешало исследователям описывать новые типы близких к эукариотам архей по метагеномным данным, и теперь их объединяют в группу Asgard.
И все же через несколько лет, в 2019 году, один из непокорных родственников эукариот был выращен японскими учеными в виде культуры. О своем успехе они сообщили в Nature в 2020 году [2]. Архея, которую с большими ухищрениями удалось вырастить исследователям, получила название Candidatus Prometheoarchaeum syntrophicum (в микробиологии принято латинские названия недавно описанных микроорганизмов, для которых еще не получены типовые штаммы, предварять словом Candidatus). О подробностях этого поистине героического исследования, занявшего в общей сложности 12 лет, можно прочитать в нашей статье «Собственной персоной: ученым удалось вырастить в лаборатории вероятного предка эукариот» [3]. Здесь стоит отметить, что Prometheoarchaeum syntrophicum оказался способен расти только в присутствии нескольких микроорганизмов, тесно связанных с ним метаболически, то есть является синтрофом (эта особенность асгардархеи и дала ей видовой эпитет). Шарообразные клетки P. syntrophicum образуют длинные разветвленные протрузии (мембранные выросты), благодаря которым могут взаимодействовать со своими партнерами по метаболизму. Вероятно, одним из микроорганизмов, вступившим в тесную дружбу с близким родственником P. syntrophicum, оказалась бактерия, которая в дальнейшем стала митохондрией, после того как архея полностью ее поглотила. Хотя в геноме P. syntrophicum нашлись гены, кодирующие сигнатурные белки эукариот, оставалось непонятным, отражается ли как-то их присутствие на внутренней организации клетки археи. Наличие сигнатурных белков эукариот указывает, что локиархеи вполне могут обладать цитоскелетом или системой внутренних мембран. Но так ли это?
К сожалению, первооткрыватели P. syntrophicum ничего не сообщили об особенностях внутренней организации археи. Однако ученые из Австрии, Швейцарии и Германии повторили успех японских коллег и вырастили в культуре асгардархею, которую назвали Candidatus Lokiarchaeum ossiferum, и детально изучили архитектуру клетки археи. Авторы этого исследования выяснили, что новооткрытая локиархея обладает мощным цитоскелетом, однако, по всей видимости, не имеет сложной системы внутренних мембран. Результаты этого исследования тоже были опубликованы в Nature в конце 2023 года [4].
Новая асгардархея была выделена из донных осадков, взятых из эстуария, который располагается на территории Словении и сообщается со Средиземным морем. Анализ пробы показал, что среди всех 16S рРНК около 4% приходятся на долю асгардархей. Образцы донного осадка были помещены в среды с различным составом, и за процессом культивирования, точнее, наращивания численности локиархей в культуре, ученые следили с помощью количественной ПЦР. Наконец, о чудо!, через целых 140 дней в одной из сред удалось выявить рост численности локиархей, однако спустя два пересева культуры ее рост остановился. Спасти ситуацию удалось с помощью той же среды, в которой вырастили локиархею P. syntrophicum. Дальнейшее усовершенствование состава среды позволило добиться того, что даже спустя несколько пересевов численность локиархей в культуре колебалась от 25% до 80% в зависимости от вариаций в некоторых ингредиентах среды.
Впрочем, помимо локиархей, с ними припеваючи жило целое сообщество микроорганизмов, включающее сульфатредуцирующую бактерию Desulfovibrio, метаногена Methanogenium, а также представителей родов Halodesulfovibrio и Methanofastidiosales (рис. 2). К слову, Methanogenium и Halodesulfovibrio входят и в число синтрофных партнеров P. syntrophicum. Как читатель, наверное, догадался, по образу жизни новая асгардархея Lokiarchaeum ossiferum оказалась очень похожа на своего «коллегу» и тоже является синтрофом, то есть зависит в метаболическом плане от других микроорганизмов, с которыми обитает в близком контакте. Примечательно, что растет она существенно быстрее, чем P. syntrophicum, и достигает большей плотности клеток в культуре, поэтому для получения культуры L. ossiferum не потребовалось нескольких лет.

Рисунок 2. Визуализация микробного сообщества, собравшегося вокруг L. ossiferum. ДНК окрашена голубым, с помощью флуоресцентной гибридизации in situ (FISH) окрашены клетки локиархей (красный), бактерий (зеленый) и представители порядка Methanomicrobiales (пурпурный).
На тот момент, когда была получена культура асгардархеи, ученые ничего не знали про то, что именно за вид им удалось вырастить. Это потом он получил название Lokiarchaeum ossiferum, а пока, чтобы понять, что же за локиархея была выращена, ученые тщательно отсеквенировали и проаннотировали ее геном. Геном новооткрытой локиархеи, временно обозначенной как Loki-B35, насчитывает чуть больше 6 миллионов пар оснований и кодирует 5119 предсказанных белков, три рРНК и 34 тРНК. Примечательно, что две из трех рРНК в этом геноме находятся в составе двух оперонов, при том, что известны геномы асгардархей, у которых есть и один, и три оперона рРНК. Авторы работы полагают, что вариабельность числа оперонов, кодирующих рРНК, у асгардархей можно рассматривать как приспособление для адаптации к меняющимся условиям окружающей среды. Стоит также отметить, что по размеру генома новая архея значительно обходит P. syntrophicum; кроме того, из предсказанных в новом геноме белков 2256 являются уникальными. 258 белков, закодированных в геноме Loki-B35, были отнесены к заветным эукариотическим сигнатурным белкам, причем у Loki-B35, в отличие от P. syntrophicum, среди этих сигнатурных белков существенно больше тех, что вовлечены в белковый транспорт и перестройку мембран. Филогенетический анализ генома Loki-B35 и геномов других архей группы Asgard показал, что Loki-B35 является родственником P. syntrophicum и ряда других локиархей, однако достаточно не похож на них, чтобы быть выделенным в отдельный род и вид. Так, вторая в истории асгардархея, выращенная в культуре, получила название Lokiarchaeum ossiferum.
Геном, конечно, позволяет узнать многое о его обладателе. Тем не менее, для всестороннего изучения организма необходимо видеть, так сказать, товар лицом. Поэтому, получив из генома L. ossiferum максимально подробные сведения, авторы работы взялись за исследование самих клеток новой локиархеи. Они имеют шаровидную форму и достигают в диаметре от 0,3 до 1 мкм, кроме того, несут многочисленные протрузии. Чтобы понять, как устроены клетки L. ossiferum внутри, ученые воспользовались мощным методом криоэлектронной томографии. Однако культура L. ossiferum, как мы помним, на самом деле представляла собой не чистую культуру асгардархеи, а сложное микробное сообщество. Следовательно, для анализа клеток L. ossiferum с помощью криоэлектронной томографии необходимо каким-то образом научиться отличать их от других микробных клеток, с которыми архея тесно связана метаболически. Более того, крайне желательно использовать для этой цели метод, не требующий обработки препарата способами, которые могут исказить внутреннюю организацию клеток, вроде химической фиксации, дегидратации или высоких температур. Авторы исследования нашли довольно изящное решение проблемы. Они предложили идентифицировать клетки L. ossiferum на томограммах с помощью структурных особенностей их рибосом, а именно, двух сегментов в составе рРНК, которые отсутствуют у бактерий и других архей. В частности, нет их и у микробов-партнеров, которые присутствуют в культуре L. ossiferum.
Как оказалось, клетки L. ossiferum имеют схожую морфологию с P. syntrophicum: в обоих случаях клетки шаровидной формы и несут длинные протрузии. Получив базовые представления о внешнем облике L. ossiferum, ученые приступили к исследованию поверхности клеток с помощью сканирующей электронной микроскопии. Применение этого метода в комбинации с криоэлектронной томографией позволило установить, что, хотя клетки L. ossiferum несут протрузии, как и у P. syntrophicum, в первом случае они имеют более вариабельную форму, часто ветвятся и могут быть с продолговатыми расширениями. Некоторые протрузии даже, наподобие мостиков, соединяли шарообразные тела нескольких клеток — к слову, подобные мостики были описаны и у других архей. Сами же клеточные покровы представлены однослойной мембраной, испещренной и покрытой множеством белков. Выглядит клеточная оболочка у L. ossiferum и вправду необычно: поверхность клеточной мембраны густо покрыта белковыми структурами, а во многих случаях она еще и подстилается плотными скоплениями белков. Эти скопления могут участвовать в организации сети протрузий, а порой они располагаются в местах большого искривления мембраны. Внутренняя, цитоплазматическая сторона мембраны тоже оказалась не без причуд. Иногда на ней удавалось идентифицировать расположенные в ряд хеморецепторы (к слову, и в геноме L. ossiferum были найдены белки, участвующие в хемотаксисе). В целом, совершенно уникальная клеточная оболочка L. ossiferum сохраняет свою необычную структуру по всей площади, в том числе и в тех местах, где протрузии отходят от тела клетки (рис. 3).

Рисунок 3. Морфология и внутреннее строение клеток L. ossiferum.
(а), (б) — внешний вид клеток, полученный с использованием сканирующей электронной микроскопии; масштабная линейка 500 нм;
(в), (д) — срезы клеток, визуализированные с помощью криоэлектронной томографии;
(г), (е) — реконструкции частей клетки в 3D, построенные с помощью нейронной сети.
На изображениях (в) и (г) — тела клеток, на (д) и (е) — протрузии. Оранжевые стрелки указывают на цитоскелетные филаменты, голубые — на сложные поверхностные структуры. На рисунках (в) и (г) масштабная линейка — 100 нм (томограмма) и 1 мкм (обзорное изображение в 2D).
Внутри клеток L. ossiferum удалось визуализировать многочисленные тонкие, порой изогнутые филаменты — наличие цитоскелета у этой археи налицо. Интересно, что филаменты часто заходят и в протрузии, а не находятся исключительно в теле клетки. А вот мембранных внутриклеточных компартментов, за исключением только двух случаев, найти в клетках L. ossiferum не удалось. По всей видимости, внутриклеточная мембранная компартментализация — не самое сильное место новой локиархеи.
Цитоскелетные филаменты L. ossiferum, попавшие на томограммы, как оказалось, состоят из двух нитей, формирующих спираль. Параметры этой двойной спирали оказались весьма близки к филаментам F-актина эукариот, а также кренактина, который имеют археи из группы Crenarchaeota. Но есть ли у локиархей белки, родственные актину или кренактину? В геноме L. ossiferum, действительно, закодированы гомологи эукариотического актина, причем сразу четыре. Филогенетический анализ показал, что один из этих гомологов попадает в число локиактинов, которые формируют сестринскую группу по отношению к эукариотическому актину и большинству ARP (актин-родственных белков). Остальные три гомолога родственны другим ARP эукариот и асгардархей. Похоже, актины локиархей прошли весьма витиеватый эволюционный путь, который сопровождался дупликацией, утратой и приобретением генов. Важно отметить, что локиактины есть во всех группах архей, входящих в Asgard, и можно предположить, что последний общий предок эукариотоподобных архей уже имел локиактин!
Используя окрашивание антителами к локиактину, авторы исследования наглядно показали, что цитоскелетные филаменты L. ossiferum состоят именно из локиактина (рис. 3). Примечательно, что в геноме L. ossiferum закодированы и белки, отвечающие за перестройки актинового цитоскелета, в частности, влияющие на полимеризацию и динамику филаментов, — эти белки гомологичны белкам гельзолину и профилину, перестраивающими сети актина у эукариот. Таким образом, можно заключить, что у L. ossiferum есть самый настоящий актиновый цитоскелет, который может претерпевать динамические изменения под влиянием актин-связывающих белков.

Рисунок 4. Визуализация филаментов локиактина в клетках L. ossiferum с помощью антител и микроскопии STED. Геномная ДНК визуализирована с помощью конфокальной микроскопии. Масштабная линейка — 1 мкм.
Итак, мы убедились, что у локиархеи L. ossiferum, родственника нашего вероятного непосредственного предка, есть не только развитая сеть разветвленных клеточных выростов, но и сложный актиновый цитоскелет. (На всякий случай уточним, что, хотя предок эукариот, скорее всего, был очень близок L. ossiferum, сама асгардархея L. ossiferum нашим с вами далеким предком не является.) Указанные особенности цитоскелета отчасти объясняют, почему археи Asgard обитают преимущественно в донных отложениях. В самом деле, наличие длинных тонких выростов делает клетку очень хрупкой, и в толще воды ее участь была бы незавидна. Устройство клеточных покровов у L. ossiferum очень необычно для архей. Чаще всего у архей поверх мембраны или, при наличии, слоя псевдопептидогликана залегает белковый S-слой, но у L. ossiferum такого слоя нет, зато поверхность клетки густо покрыта множеством белков, не формирующих единого слоя. Можно предположить, что именно поверхностные белки дали L. ossiferum возможность формировать контакты с другими клетками и тем самым заложили основу для эукариогенеза. Наконец, развитый цитоскелет может обеспечивать локиархее необходимый уровень компартментализации внутриклеточных процессов даже в отсутствие мембранных органелл. Кроме того, актиновый цитоскелет может быть основой для многих процессов, в которых задействованы эукариотические сигнатурные белки, например, для транспортировки макромолекул по клетке. А наличие L. ossiferum в лабораторной культуре делает возможным непосредственное изучение всех захватывающих процессов, связанных с происхождением эукариотической клетки.
Литература
- Anja Spang, Jimmy H. Saw, Steffen L. Jørgensen, Katarzyna Zaremba-Niedzwiedzka, Joran Martijn, et. al.. (2015). Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature. 521, 173-179;
- Hiroyuki Imachi, Masaru K. Nobu, Nozomi Nakahara, Yuki Morono, Miyuki Ogawara, et. al.. (2020). Isolation of an archaeon at the prokaryote–eukaryote interface. Nature. 577, 519-525;
- Собственной персоной: ученым удалось вырастить в лаборатории вероятного предка эукариот;
- Thiago Rodrigues-Oliveira, Florian Wollweber, Rafael I. Ponce-Toledo, Jingwei Xu, Simon K.-M. R. Rittmann, et. al.. (2023). Actin cytoskeleton and complex cell architecture in an Asgard archaeon. Nature. 613, 332-339.
Комментарии


