Транскрипция в хроматине: как проходить сквозь стены
24 марта 2015
Транскрипция в хроматине: как проходить сквозь стены
- 3821
- 0
- 8
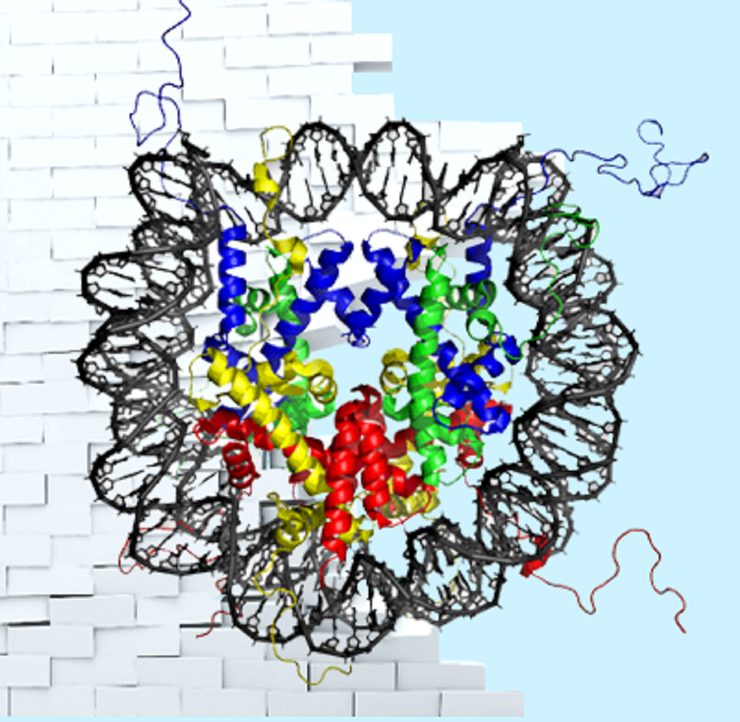
Нуклеосома состоит из гистонов и связанной с ними ДНК, совершающей 1,65 оборота. В центре белкового ядра расположен тетрамер из гистонов (Н3-Н4)2, фланкированный с двух сторон димерами гистонов Н2А-Н2В.
Рисунок в полном размере.
изображение с сайта www.molecularstation.com
-
Автор
-
Редакторы
Перед РНК-полимеразой стоит сложная задача реализации генетической информации путем осуществления транскрипции. На своем пути фермент встречает массу препятствий. ДНК находится в комплексе с белками и плотно упакована, образуя нуклеосомы, которые создают барьер для полимераз, но несут важные регуляторные сигналы. Так как же транскрипционная машина проходит через нуклеосомы?
Гистоновый код
Молекула ДНК упакована в ядре плотно. Она, объединяясь с белками-гистонами, образует хроматин, который имеет определенную структуру: его единицей является нуклеосома, состоящая из восьми молекул гистонов и намотанной на них ДНК (примерно 160 п.н.). Дополнительный девятый (линкерный) гистон H1 не входит в центральное нуклеосомное «ядро», а фиксирует нить ДНК на поверхности каждой нуклеосомы.
Хроматин находится под чуткой регуляцией многих систем. Он начинает строиться сразу, как только перед делением клетка удваивает свой генетический материал [1]. Изучение его структуры важно не только с теоретической точки зрения: известно, что в раковых клетках нарушено строение хромосом, увеличено ядрышко (немембранное образование в ядре клетки, легко различимое с помощью микроскопии), и вскрытие причин этих аномалий позволит разработать новые терапевтические подходы. Интересно, что с изменением размера ядрышка связывают колебания активности белков, облегчающих транскрипцию в хроматине, — например, белкового комплекса FACT [2]. О преодолении сложностей транскрипции нуклеосомной ДНК и пойдет речь дальше.
Нуклеосомная ДНК 1,65 раза оборачивается вокруг белкового ядра нуклеосомы, которое состоит из гетеродимеров гистонов Н2А, Н2В, Н3 и Н4 [3]. N-концы гистонов выходят за пределы такой «катушки» (видно на заглавном рисунке) и играют важную регуляторную роль. На них, как флажки, появляются различные модификации, которые дают сигналы другим молекулам. Набор модификаций представляет собой часть эпигенетического кода — гистоновый код, который участвует в регуляции экспрессии генов. Выделяют сигналы, способствующие транскрипции определенных генов, или, наоборот, препятствующие ей. Некоторые модификации говорят о том, что ДНК в этой зоне нуждается в ремонте (репарации), или о том, что здесь закодирован элемент, который не несет информации о белке, а является регуляторным. Модификации гистонов узнаются, влекут за собой каскады реакций, по необходимости убираются или заменяются на новые. Таким образом, гистоновый код динамичен: он отражает потребности клетки в конкретный момент времени и очень важен для ее нормальной жизнедеятельности.
Нуклеосомы преграждают путь РНК-полимеразам
РНК-полимеразы реализуют генетическую информацию, синтезируя РНК на матрице ДНК. Прочитывая ген, они должны получить доступ к каждому нуклеотиду цепи. Однако когда нуклеотидная цепочка находится в составе нуклеосомы, это затрудняет работу полимеразы. Гистоны связывают те участки ДНК, которые должны быть прочитаны. Для преодоления такого рода препятствия РНК-полимеразы могут «скинуть» гистоны со своего пути, но тогда потеряются и белковые модификации, под регуляцией которых находился ген. Однако существуют и способы преодоления нуклеосом с их сохранением на ДНК. На данный момент описаны оба варианта и, видимо, они сосуществуют в клетке.
Полимераза прошла, нуклеосома осталась, но изменилась

Рисунок 1. U-образная форма нуклеосом в районе активных генов.
трехмерная реконструкция из [5]
Клетки одного организма содержат одинаковый набор генов, но при этом они разные: есть нервные, есть мышечные и т.д. В определенном типе клеток активны характерные для него гены. РНК-полимераза прочитывает только их, остальные заблокированы и молчат. Нуклеосомы активных генов несут специальные модификации (например, ацетилирование гистонов), есть у них и другие особенности [4–6]. Нуклеосомы в тех местах, где проходит РНК-полимераза, часто имеют U-образную форму (рис. 1).
Нулеосома в районе активных генов может вообще «раскрываться». То есть ядро из восьми гистонов делится на две части из четырех молекул. «Развернутые» нуклеосомы накапливаются ближе к концу гена [7]. Это может быть обусловлено тем, что РНК-полимераза, расплетая двойную спираль ДНК, создает в ней напряжение. Особенно сильным оно будет в конце гена. Интересно, что после прекращения транскрипции обычная форма нуклеосом восстанавливается [8].
РНК-полимеразы, объединяясь, сметают нуклеосомы
Гены могут быть активны по-разному: с некоторых РНК считывается относительно редко, с некоторых гораздо чаще. В том числе существуют гены, работающие только в определенных тканях. Например, в 85% типов рака активен ген теломеразы — особого белка, который способен достраивать концы хромосом. Это один из механизмов, помогающих клеткам делиться бесконечно при патологии [9].
В случае интенсивной транскрипции гистоны могут терять связь с ДНК, если РНК-полимеразы прочитывают ген одна за другой. Уже после прохода через нуклеосому первой полимеразы часто пара гистонов Н2А–Н2В уходит в раствор (рис. 2), но шанс «выжить» у оставшихся шести гистонов еще есть. А при активной транскрипции, когда две РНК-полимеразы идут прямо друг за другом, с ДНК уходит весь октамер [10]. Интересно, что такие следующие друг за другом ферменты работают более эффективно. Они лучше справляются с тем барьером, который создает нуклеосома. Вторая РНК-полимераза как бы подталкивает первую и не дает ей отступать назад (что иногда случается).

Рисунок 2. Во время транскрипции гистоны могут уходить из состава нуклеосомы. Если на участке ДНК проходит только один раунд транскрипции, то теряются преимущественно гистоны Н2А–Н2В. Если РНК-полимеразы идут друг за другом, то ДНК может полностью высвобождаться.
адаптировано из [10]
РНК-полимеразы могут пройти через нуклеосому
Итак, если уровень транскрипции не очень высок, то у нуклеосом есть шанс «выжить». Это влечет за собой возможность сохранения гистоновых модификаций в том же месте хроматина, где они и были раньше, что может быть важно для жизни клетки. Американскими учеными с российскими корнями в 2009 году был предложен механизм прохождения через нуклеосому без ее удаления для РНК-полимеразы II, транскрибирующей большинство генов эукариот [11].
Постулируется, что нуклеосома сохранится, если при расплетении нуклеотидной цепи в каждый момент времени гистоны будут связывать хотя бы часть нуклеосомной ДНК. Как это возможно, если РНК-полимераза должна прочитать все «буквы» без исключения? Оказывается, важнейшим моментом такого механизма является заключение фермента в петлю. То есть возможно перехватывание гистонов цепочкой ДНК. Опытным путем показали, что сначала РНК-полимераза расплетает только часть нуклеосомной ДНК. Затем, когда фермент прочитывает эту свободную от связей с белками часть цепи, она закручивается обратно. В этот момент восстанавливаются связи с гистонами у той ДНК, что уже оказалась позади РНК-полимеразы. Такое состояние в литературе называют нулевой петлей. Затем фермент продолжает свой путь и освобождает от белков вторую половину нуклеосомной ДНК (рис. 3). Важно отметить, что при такой работе системы некоторые гистоны все-таки теряются. Часто из состава нуклеосомы уходит один гетеродимер белков Н2А и Н2В.

Рисунок 3. Механизм «сквозной» транскрипции. 1) РНК-полимераза приближается к нуклеосоме. 2) Фермент «отворачивает» часть нуклеосомной ДНК, освобождая ее от связи с гистонами. 3) РНК-полимераза оказывается в петле, когда восстанавливаются связи ДНК, оставшейся позади фермента, с гистонами. 4) Вторая часть нуклеосомной ДНК расплетается. 5) Когда РНК-полимераза заканчивает транскрипцию нуклеосомной ДНК, последняя восстанавливает свою конформацию.
Итак, нуклеосомы не только помогают компактно уложить цепь ДНК, но и несут важную эпигенетическую информацию. Оказывается, что после транскрипции они могут как пропадать, так и оставаться на прежнем месте. Нуклеосомы могут изменять свою форму, а затем восстанавливать прежнюю. Их судьба в таких процессах, как транскрипция или репликация, представляет большой интерес для науки. Изучение эпигенетики важно для медицины, для понимания развития живых организмов, их поведения и регуляции генома в целом [12]. Накопленные данные уже позволили получить ряд лекарственных препаратов. Они влияют на белки, создающие или убирающие эпигенетические сигналы. Например, такие лекарства используют для лечения лейкозов и лимфом [13].
Литература
- Катится, катится к ДНК гистон;
- Размер имеет значение;
- Новые возможности в лечении наследственных заболеваний крови;
- Genoveva A. Nacheva, Dmitrii Y. Guschin, Olga V. Preobrazhenskaya, Vadim L. Karpov, Konstantin K. Ebralidse, Andrei D. Mirzabekov. (1989). Change in the pattern of histone binding to DNA upon transcriptional activation. Cell. 58, 27-36;
- D. Bazett-Jones. (1996). Visualization and analysis of unfolded nucleosomes associated with transcribing chromatin. Nucleic Acids Research. 24, 321-329;
- Как «вырезать» вирус?;
- Aurélien Bancaud, Gaudeline Wagner, Natalia Conde e Silva, Christophe Lavelle, Hua Wong, et. al.. (2007). Nucleosome Chiral Transition under Positive Torsional Stress in Single Chromatin Fibers. Molecular Cell. 27, 135-147;
- Richard Sterner, Lidia C. Boffa, Thelma A. Chen, Vincent G. Allfrey. (1987). Cell cycle-dependent changes in conformation and composition of nucleosomes containing human histone gene sequences. Nucl Acids Res. 15, 4375-4391;
- Теломеры и новые мишени протоонкогенной терапии;
- Olga I. Kulaeva, Fu-Kai Hsieh, Vasily M. Studitsky. (2010). RNA polymerase complexes cooperate to relieve the nucleosomal barrier and evict histones. Proc. Natl. Acad. Sci. U.S.A.. 107, 11325-11330;
- Olga I Kulaeva, Daria A Gaykalova, Nikolai A Pestov, Viktor V Golovastov, Dmitry G Vassylyev, et. al.. (2009). Mechanism of chromatin remodeling and recovery during passage of RNA polymerase II. Nat Struct Mol Biol. 16, 1272-1278;
- Эпигенетика поведения: как бабушкин опыт отражается на ваших генах?;
- Пилюли для эпигенома.



Комментарии
0Чтобы оставить комментарий, необходимо
войти