Как составлялся геном эукариот: эндосимбиоз VS. непрерывный горизонтальный перенос
22 сентября 2015
Как составлялся геном эукариот: эндосимбиоз VS. непрерывный горизонтальный перенос
- 4067
- 0
- 14
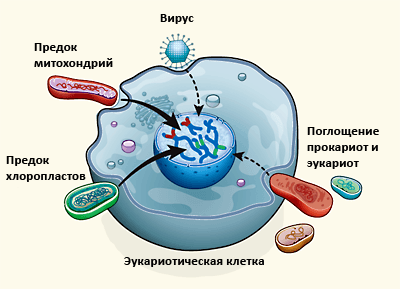
Ядерный геном эукариот отличается огромным разнообразием генов, полученных в том числе и от бактерий с археями. Чужеродная ДНК может попасть в клетку двумя способами: с вирусами и при поглощении других клеток. Пунктирными стрелками обозначены пути приобретения генов, которые, по мнению авторов статьи [7], оказали минимальное воздействие на геном эукариот.
[7], рисунок адаптирован
-
Автор
-
Редакторы
Результаты работы крупной международной группы ученых, опубликованные в августовском Nature, добавили огоньку в давнее противостояние двух точек зрения на появление в эукариотическом геноме генов прокариот. Одна концепция утверждает, что прокариотические гены попали в ядерные клетки главным образом вместе с предками органоидов, другая — что горизонтальный перенос генов в мире эукариот — процесс распространенный и непрерывный. Новая работа подтачивает фундамент второй концепции, приводя альтернативные объяснения очевидной неоднородности распределения генов у эукариот.
Ядерный геном эукариот — это своеобразная мозаика из ДНК, приобретенной из различных источников. Например, из митохондрий и хлоропластов — эукариотических органоидов, которые произошли от прокариотических клеток, ассимилированных в процессе эндосимбиоза. Хотя эти органоиды и сохраняют часть своей первоначальной ДНК, внушительное количество их наследственного материала переместилось в ядро хозяина вследствие эндосимбиотического переноса генов [1]. В случае с хлоропластами дальнейшие эукариот-эукариотические эндосимбиозы распространили эти органоиды и связанные с ними гены по ветвям эволюционного дерева [2].
У прокариот вертикальное наследование (клональный рост) дополняется процессом рекомбинации в результате горизонтального переноса генов (ГПГ) , осуществляемого путем трансформации, конъюгации, трансдукции. Эти механизмы работают однонаправленно — от донора к реципиенту — и приводят к формированию пангеномов [3].
О горизонтальном генетическом переносе у прокариот и многообразии его виновников — мобильных генетических элементов — рассказывает статья «Мобильные генетические элементы прокариот: стратификация „общества“ бродяжек и домоседов» [4].
Для эукариот характерна половая рекомбинация — процесс двунаправленный, обоюдный, так как перекраивается и отцовская, и материнская ДНК. Наследование же происходит почти исключительно вертикально из-за дефицита систем ГПГ, подобных прокариотическим. Отделение половых клеток от остальной части организма у многоклеточных эукариот, как полагают, служит прочным барьером для интродукции чужеродной ДНК. Однако это предположение многие исследователи считают сомнительным и приводят примеры горизонтального переноса генов и у животных, и у растений:
- вирусы могут служить векторами для переноса генов и облегчать ГПГ у эукариот так же, как у прокариот [5];
- одноклеточные эукариоты могут приобретать чужеродную ДНК, просто поглощая и переваривая добычу.
Никто не отрицает эпизод мощного вброса прокариотических генов при «одомашнивании» хлоропластов и митохондрий, когда часть наследственного материала эндосимбионтов переместилась в ядро клетки [6]. Однако большинство случаев горизонтального переноса среди эукариот было выявлено исключительно на основе филогенетических несоответствий — обнаружения генов, эволюционные истории которых не согласуются с известными или предсказанными взаимоотношениями организмов. Здесь следует учесть еще и то, что при построении деревьев часто возникают артефакты, отчего страдает воспроизводимость результатов , разнятся их интерпретации [7].
Проблемы с воспроизводимостью и достоверностью результатов биологических, медицинских, психологических исследований довели-таки научное сообщество и спонсоров до отчаяния — до тайных собраний, отказа от традиционных критериев, массового освоения интернет-облаков и генерации прожектов на тему открытой и честной науки: «Мечты о воспроизводимости» [8].
Еще более спорное утверждение — об обильном и постоянном горизонтальном переносе генов от прокариот к эукариотам [9]. Если это верно, то следствием таких процессов должен быть кумулятивный эффект в эволюции эукариот, который должен обнаруживаться при анализе большого числа геномов разных эукариотических линий. Эта гипотеза противопоставляется теории эндосимбиоза, которая утверждает, что приобретение чужеродных генов эукариотами соответствует времени происхождения органоидов и вторичного эндосимбиоза у водорослей (рис. 1).

Рисунок 1. Эндосимбиотическая эволюция. Внутриклеточные эндосимбионты, которые первоначально произошли от свободноживущих прокариот, сыграли важную роль в эукариотической эволюции, породив два цитоплазматических органоида. Митохондрии развились из α-протеобактерий, а хлоропласты — из цианобактерий. Оба органоида внесли весомый вклад в совокупный генόм эукариотических ядер. На рисунке показана схема эволюции эукариот с акцентом на моментах включения митохондрий и хлоропластов в эукариотическую линию посредством эндосимбиоза и дальнейшей параллельной эволюции ядерного и органоидных геномов. Клетка-хозяин, которая приобрела пластиды, вероятно, обладала двумя жгутиками. Общепринято, что хозяин, заполучивший митохондрии (внизу справа), состоял в родстве с археями, однако о характере его внутриклеточной организации (прокариотической, эукариотической или промежуточной), возрасте, биохимии и генном составе биологи пока договориться не могут. Полагают, что хозяин был одноклеточным.
[2], рисунок адаптирован
Основание для заявлений о широком распространении горизонтального переноса генов у эукариот возникло после выявления неожиданных ветвей филогенетических деревьев и неоднородного распределения генов между линиями. Действительно, некоторые случаи линия-специфичного горизонтального переноса генов в эукариотические клетки неопровержимы: это включения бактериальных генов в хромосомы насекомых и вирусные вставки в процессе эволюции плацентарных [10], [11]. И все же, если ГПГ у эукариот — непрерывный процесс, он должен привести к кумулятивному эффекту. Даже при очень низкой средней частоте переноса чужой ДНК в геном эукариот (например, 0,5%) приобретенные гены будут наследоваться потомками, которые продолжат приобретать новые гены. В итоге должны появляться все более и более различающиеся генетические линии эукариот, которые будут со временем расходиться все дальше друг от друга. Как раз такую схему эволюции мы наблюдаем в прокариотическом мире, где работает в полную силу ГПГ и накапливаются пангеномы.
Так как же все-таки объяснить присутствие прокариотических генов в хромосомах эукариот — постепенным их накоплением в результате ГПГ или эпизодическим заимствованием от органоидов? Международная группа ученых из Германии, Новой Зеландии, Израиля, Португалии и других стран попыталась тщательно проверить обе гипотезы [3].
Распределение генов подтверждает теорию эндосимбиоза
В исследовании использовали около миллиона белковых последовательностей 55 эукариот шести супергрупп (рис. 2) и более шести миллионов последовательностей прокариот (300 000 от 134 архей, остальные — от 1847 бактерий). В результате выявили 2585 обособленных семейств белковых последовательностей, каждое из которых включало связанные (гомологичные) белки, кодируемые двумя или более эукариотическими и пятью или более прокариотическими геномами: эти семейства были названы эукариот-прокариотическими кластерами(ЭПК). Остальные — эукариотспецифичными кластерами (ЭСК).

Рисунок 2. Эволюция эукариотического генома. Внизу показаны 6 супергрупп эукариот. Гены, унаследованные от эукариотического предка и митохондрий, обозначены пшеничным цветом, гены пластидного происхождения — зеленым (у организмов со вторичными пластидами не обозначены). Высота вертикальных прямоугольников отражает размер геномов (число ЭПК) «узловых» эукариот. Заметно, что эволюция эукариот сопровождалась потерей генов. Последовательности архейного происхождения (преимущественно «информационные»), унаследованные от линии архей, захватившей предка митохондрии и развившейся в эукариот, утрачивались реже, чем прочие гены («операционные», метаболические) [3]. Нажмите на изображение, чтобы увеличить.
[3], рисунок изменен
87% деревьев первого кластера показали эукариотическую монофилию, для 12% она не исключалась. Однако остальные вовсе не обязательно представляют собой продукты горизонтального переноса генов, поскольку: а) эндосимбиоз привносит генетическую избыточность (например, в «хозяйскую» линию проникают гены рибосом эндосимбионтов); б) у этих деревьев выявлены признаки загрязнения при секвенировании; в) иногда молекулярная филогенетика... просто ошибается. Даже если предположить, что эти деревья отражают результат ГПГ, эукариоты все равно не расширили свой генный репертуар за счет принятых последовательностей, поскольку повторно приобрели гены, уже присутствующие в эукариотической линии.
Сделав такой вывод, исследователи сфокусировались на изучении ЭПК-деревьев с эукариотической монофилией. Больше половины из этих кластеров (1060) имели прямое отношение к внедрению фотосинтеза в эукариотическую линию и его распространению путем вторичного эндосимбиоза (при формировании красных вторичных пластид страменопил, альвеолят, Guillardia, Emiliania и зеленых вторичных пластид Bigelowiella) [12], [13].
В той же группе (ЭПК) лишь единичные прокариотические кластеры были монофилетическими, что отражает ситуацию с пангеномами и распространенным среди прокариот горизонтальным переносом генов. Если бы эукариоты были вовлечены в пангеномный перенос генов вместе с прокариотами, они бы тоже обладали пангеномами, чего не наблюдается. Это наводит на мысль, что, за исключением эпизодов «приручения» предков митохондрий и пластид, эукариоты были изолированы от ГПГ, ведущего к формированию пангеномов у прокариот.
Эволюция эукариотических генов вертикальна
Существует только два механизма, которые могли привести к образованию 87% монофилетических деревьев в группе ЭПК:
- ген белка присутствовал в общем предке и вертикально наследовался потомками;
- ген был приобретен одним из членов группы и затем распространен посредством эукариот-эукариотического ГПГ.
В первом случае деревья на всем своем протяжении будут сходны с деревьями ЭСК. Во втором они будут отличны друг от друга и от каждого следующего вновь возникшего.
Ученые показали, что эволюция эукариотических генов «вопиюще» вертикальна, в то время как альтернативный механизм, связанный с эукариот-эукариотическим горизонтальным переносом генов, полностью исключили для всех шести супергрупп. В качестве доказательства они привели пример с супергруппой архепластид (растений в широком смысле). Анализ ЭПК-деревьев показал, что:
- хотя совсем немного архепластидспецифичных ЭПК можно отследить прямо до цианобактерий, все архепластидспецифичные ЭПК представлены у общего предка Archaeplastida;
- появление этих ЭПК совпадает с образованием пластид;
- многие кластеры имеют непосредственное отношение к фотосинтезу;
- но: их сестринские прокариотические группы филогенетически неоднородны, то есть гомологичные гены есть у разных прокариот (не только у цианобактерий).
Последнее наблюдение можно объяснить двумя способами.
- Если считать эти прокариотические таксоны биологическими донорами, то тогда в происхождение пластид должны быть вовлечены сотни независимых горизонтальных переносов генов от сотен различных доноров. То есть фотосинтезирующие эукариоты должны были строиться ген за геном, при этом исключались всякие мутации, инактивирующие перенесенные гены до сборки пластиды в готовую функциональную единицу.
- Деревья вводят в заблуждение, и архепластидспецифичные ЭПК приобретены от предка пластид, уже имеющего функционирующий фотосинтетический аппарат, который нужно было просто интегрировать в эукариотическую линию посредством переноса необходимых генов из органеллы в ядро (эндосимбиотического переноса генов).
Второй вариант, безусловно, более правдоподобен (рис. 3).
Эпизодический приток и избирательная потеря генов
О митохондриях можно рассуждать по аналогии с пластидами. Хозяин пластид изначально был гетеротрофом, а переход к автотрофии произошел посредством эндосимбиоза с переносом генов. Распределение генов, полученное в новом исследовании, это доказывает. Хозяевами митохондрий на каком-то этапе стали археи , а эндосимбиоз с переносом генов из органеллы в ядро хозяина привел к возникновению хемиосмотического синтеза АТФ. Как и пластиды, митохондрии не могли быть сконструированы пошагово, путем последовательных горизонтальных переносов генов: сотни беспорядочно приобретенных для создания дыхательного органоида генов не могли поддерживаться в клетке очищающим отбором до момента сборки полноценной митохондрии. Перенести необходимые гены в ядро из дышащего эндосимбионта было бы несравнимо проще (рис. 3).
Особенности эволюции архей, насыщенной судьбоносными заимствованиями генов у бактерий, подробно разобраны в статье «Закинули археи эволюционный невод и вытянули...» [14].

Рисунок 3. Сеть эндосимбиотического переноса генов (ЭПГ). На карту референсной филогении нанесена сеть, соединяющая предполагаемых доноров генов с общими предками архепластид и всех эукариот. Интенсивность серых линий (от белого до черного) в прокариотических референсных деревьях показывает, насколько часто конкретная ветвь «перекрывалась» дендрограммами отдельных генов, использованных для формирования итоговой топологии. Цветные линии показывают приток генов в точках происхождения эукариот и образования пластид. Цвет линий (по шкале) отражает, сколько общих генов обнаружено у эукариот с тем или иным прокариотическим таксоном (т.е. возможный донорский вклад конкретной группы бактерий или архей). Вторичные ЭПГ обозначены изолированными молочно-зеленой и молочно-красной линиями. Нажмите на изображение, чтобы увеличить.
Но как объяснить неоднородное распределение генов между линиями эукариот, о котором говорилось в начале статьи? У прокариот такое положение дел указывает на ГПГ (за исключением изолированных видов с редукционной эволюцией), что подталкивает к подобному выводу и в отношении эукариот. Однако, как показали ученые, вертикальное наследование у эукариот характеризуется дифференциальной (избирательной) потерей генов, которая наилучшим образом объясняет неоднородное распределение генетической информации в этом домене. Это справедливо как для ЭПК, так и для ЭСК. В то же время приобретения новых генов — события эпизодические, соответствующие периоду формирования эндосимбионтов.
Некоторые наборы генов из эукариот-прокариотических кластеров кажутся специфичными для конкретных линий приобретениями: например, 67 генов у диктиостелид, 160 генов у опистоконтов и 210 генов, предположительно приобретенных предком сухопутных растений. Если эти гены были заимствованы путем горизонтального переноса недавно, например, в период становления сухопутных растений 450 млн лет назад, они должны быть более сходными с их прокариотическими «родственниками», чем гены, полученные от эндосимбионтов. Однако обнаружено обратное. Гены-кандидаты, подозреваемые в геном-специфичном (горизонтальном) проникновении из про- в эукариот, тоже были выявлены, но встречаются эти предполагаемые продукты ГПГ приблизительно в 4–10 раз реже, чем ядерные вставки митохондриальных и пластидных последовательностей, а число их уменьшается по мере обновления описаний геномов.
Заключение
Анализ распределения эукариот-прокариотических кластеров выявил ожидаемо большой эволюционный след, оставленный приобретением хлоропластов. Сотни ядерных генов отсутствуют у эукариотических видов, не имеющих хлоропластов, но распространены среди различных групп эукариот-фотосинтетиков и цианобактерий, от которых хлоропласты и произошли. Эндосимбиотический след в митохондриальной эволюции тоже очевиден, хотя менее отчетлив.
Несмотря на множество сообщений о горизонтальном переносе генов из прокариот в эукариоты и между эукариотами, комплексные исследования всех деревьев не выявили никаких доказательств кумулятивного эффекта непрерывного ГПГ на эволюцию эукариотических геномов. Это указывает на одну из следующих возможностей:
- перенесенные горизонтально гены, специфичные для определенных линий, быстро элиминируются;
- восприимчивые к ГПГ линии не дают начало эволюционно стабильным потомкам (тупиковы);
- многие предполагаемые продукты ГПГ на самом деле не являются линия-специфичными, и при расширении аналитической выборки эукариот они будут обнаружены у других отдаленных родственников, что докажет причинную роль дифференциальной потери в возникновении неоднородности распределения генов;
- возможна любая комбинация описанных вариантов.
Итак, по мнению участников исследования [3], два эпизода эндосимбиотического переноса генов (один — от митохондрий, другой — от пластид) с последующей избирательной потерей ДНК объясняют филогению и неоднородное распределение бактериальных генов в эукариотах. Кроме того, авторы дали возможность оценить достоинства и недостатки разных подходов, применяемых для различения горизонтального и вертикального наследований у эукариот. Но, без сомнения, вопрос остается открытым для обсуждения.
Литература
- Lane C.E. and Archibald J.M. (2008). The eukaryotic tree of life: endosymbiosis takes its TOL. Trends. Ecol. Evol. 23, 268–275;
- Timmis J.N., Ayliffe M.A., Huang C.Y., Martin W. (2004). Endosymbiotic gene transfer: organelle genomes forge eukaryotic chromosomes. Nat. Rev. Genet. 5, 123–135;
- Ku C., Nelson-Sathi S., Roettger M., Sousa F.L., Lockhart P.J., Bryant D. et al. (2015). Endosymbiotic origin and differential loss of eukaryotic genes. Nature. 524, 427–432;
- Мобильные генетические элементы прокариот: стратификация «общества» бродяжек и домоседов;
- Вирусные геномы в системе эволюции;
- Как появились митохондрии (рассказ, похожий на сказку);
- Archibald J.M. (2015). Gene transfer in complex cells. Nature. 524, 423–424;
- Мечты о воспроизводимости;
- Huang J. (2013). Horizontal gene transfer in eukaryotes: the weak-link model. Bioessays. 35, 868–875;
- Husnik F., Nikoh N., Koga R., Ross L., Duncan R.P., Fujie M. et al. (2013). Horizontal gene transfer from diverse bacteria to an insect genome enables a tripartite nested mealybug symbiosis. Cell. 153, 1567–1578;
- Mi S., Lee X., Li X., Veldman G.M., Finnerty H., Racie L. et al. (2000). Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis. Nature. 403, 785–789;
- Gould S.B., Waller R.F., McFadden G.I. (2008). Plastid evolution. Annu. Rev. Plant. Biol. 59, 491–517;
- Элементы: «Возникновение и эволюция пластид»;
- Закинули археи эволюционный невод и вытянули....
Комментарии


