Как живет бактерия, видимая невооруженным глазом
07 апреля 2022
Как живет бактерия, видимая невооруженным глазом
- 1627
- 0
- 2
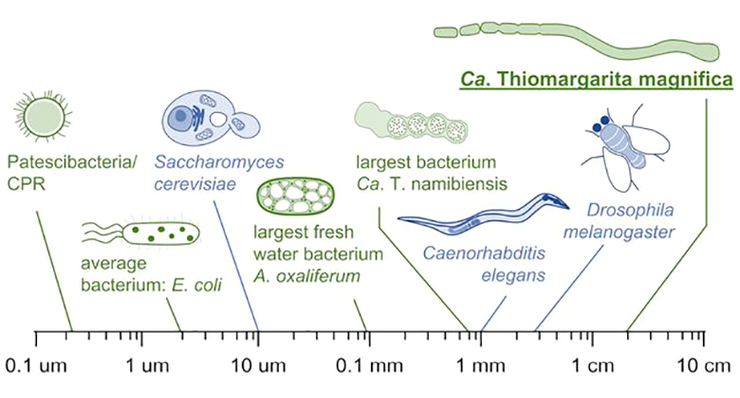
Гигантская бактерия Thiomargarita magnifica по размеру не только обходит все известные прокариотические клетки, но и даже некоторые многоклеточные эукариотические организмы, например, нематоду Caenorhabditis elegans и плодовую мушку Drosophila melanogaster.
Рисунок в полном размере.
иллюстрация из [1]
-
Автор
-
Редакторы
Темы
Бактериальные клетки, как правило, значительно меньше эукариотических и достигают в среднем 2 мкм в диаметре. Конечно, из этого правила есть любопытные исключения — некоторые бактерии имеют размер порядка 750 мкм (например, серная бактерия Thiomargarita namibiensis). Но недавно была открыта бактерия, которая по размеру превосходит не только всех ранее известных прокариот, но и большинство эукариотических клеток. Американские ученые обнаружили в мангровых зарослях Гваделупы нитевидную бактерию длиной 9 мм, причем отдельные ее клетки достигают ни много ни мало 2 см в длину. Новая бактерия получила предварительное название Thiomargarita magnifica. Как живет эта удивительная бактерия? Давайте разбираться.
Термином «микроорганизм» обычно называют одноклеточные существа, которые различимы в световой микроскоп, но не видны невооруженным глазом. К микроорганизмам относятся почти все прокариоты, а также ряд эукариот, например, дрожжи. Большинство вирусов можно рассмотреть только в электронный микроскоп, и чаще всего вирусы в понятие «микроорганизмы» не включаются. Впрочем, в последние годы и без того размытые границы микромира стали еще более условными. Открытие гигантских вирусов, поражающих амеб (в частности, мимивирусов), показало, что к микроорганизмам вполне можно отнести и некоторые вирусы (подробнее о гигантских вирусах можно прочитать в нашей статье [2]). Еще один мощный удар по концепции микроорганизмов нанесли гигантские бактерии, некоторых из которых можно рассмотреть и без всякого микроскопа.
Одним из самых ярких представителей гигантских бактерий является род Thiomargarita. Большинство его представителей населяют моря и океаны, занимаются окислением сульфида S2–, причем размер их примерно в 50 раз превышает типичные параметры бактериальных клеток. До недавнего времени самой крупной бактерией считалась Thiomargarita namibiensis, которую нашли в донных отложениях континентального шельфа Намибии. Наиболее крупные клетки этого вида достигают 750 мкм в длину (клетки такого размера можно рассмотреть даже невооруженным глазом), а большинство имеет диаметр порядка 100–300 мкм (такие клетки можно разглядеть уже с помощью бинокуляра) [3]. Однако недавно была описана новая представительница рода Thiomargarita, отдельные клетки которой достигают целых 2 см в длину! Новую бактерию, найденную в мангровых зарослях на территории Гваделупы, назвали Thiomargarita magnifica (точнее, Candidatus Thiomargarita magnifica) [1].

Рисунок 1. Морфология и ультраструктура клеток Thiomargarita magnifica.
(а) — внешнее строение клетки T. magnifica, визуализированной с помощью светового микроскопа. Перетяжки в верхней части клеток отделяют апикальные почки. Видно, что цитоплазма клетки заполнена множеством мембранных пузырьков.
(б) — внешний вид клеток T. magnifica на разных стадиях развития.
(в) — мембрана клетки T. magnifica окрашена красителем FM 1-43x. Видно, что септ внутри клетки нет, а апикальные почки отделены неполными перегородками.
(г), (д), (е) — ультраструктура клетки T. magnifica, визуализированной с помощью электронного микроскопа, где (г) — апикальный конец клетки вблизи апикальных почек. Видно, что почти все пространство внутри клетки занимает гигантская вакуоль (v), а цитоплазма оттеснена к периферии.
(д) — более крупное увеличение участка, обведенного прямоугольником на рисунке (г). Видны серные гранулы (s), вакуоль (v) и многочисленные пепины (так ученые, описавшие T. magnifica, предложили называть мембранные пузырьки, содержащие генетический материал, см. далее в тексте).
(е) — более крупное увеличение участка, обведенного прямоугольником на рисунке (д). Стрелки указывают на пепины.
Внешний вид этой бактерии довольно своеобразен. «Букеты» из филаментов, состоящих из сантиметровых нитчатых клеток T. magnifica, прикрепляются к листьям мангрового кустарника Rhizophora mangle. Сами филаменты могут достигать в длину несколько сантиметров. Впрочем, глубоководные представители рода Thiomargarita имеют похожую морфологию. На протяжении большей части своей длины клетки T. magnifica сохраняют нитевидную форму, однако у одного из концов часто находятся 1–4 маленькие почки — дочерние клетки, образующиеся в результате неравного деления (рис. 1). Филаменты T. magnifica окружены слизистым матриксом, однако, в отличие от других гигантских бактерий, на их поверхности отсутствуют эпибиотические бактерии — бактерии, которые живут во внешнем слизистом чехле более крупных бактерий (почему, мы расскажем позднее) [1].
Ученые, обнаружившие эту бактерию, поначалу были уверены, что столь крупные нитевидные клетки на самом деле являются цепочкой из нескольких клеток, разделенных септами. Каково же было их удивление, когда на электронных микрофотографиях клетки T. magnifica никаких септ обнаружено не было! Даже в области концевых почек сплошных септ нет, есть только неполные перегородки. Электронная микроскопия выявила еще одну, не менее удивительную особенность клеток T. magnifica (кажется, слово «удивительный» в отношении этой бактерии можно применять бесконечно). Выяснилось, что большая часть клетки занята гигантской вакуолью, на долю которой приходится 70–80% объема клетки! Цитоплазма при этом оттеснена на периферию и образует тоненький слой около 4 мкм толщиной. В цитоплазме были выявлены многочисленные везикулы диаметром около 2 мкм, которые, как оказалось, выполняют роль серных гранул: метаболизм T. magnifica построен на окислении сульфида до элементарной серы, которая откладывается в составе упомянутых гранул. Кроме того, в цитоплазме присутствуют и более мелкие везикулы диаметром порядка 1 мкм. Авторы, описавшие бактерию, предположили, что в этих маленьких пузырьках может находиться часть генетического материала гигантской бактерии в силу ее высокополиплоидности (об организации генетического материала T. magnifica мы подробно поговорим чуть позже) [1].
Считается, что окруженные мембраной органеллы — удел эукариотических клеток. В частности, клеточное ядро представляет собой полость, окруженную двумя мембранами и содержащую внутри геномную ДНК. Впрочем, помещение генетического материала в «мешок», образованный липидным бислоем, описано и у некоторых бактерий, таких как планктомицеты. У одного представителя группы Atribacteria была даже описана гигантская вакуоль, занимающая большую часть объема клетки и содержащая генетический материал бактерии. Но все же у подавляющего большинства бактерий геномная ДНК находится в особой зоне цитоплазмы, известной как нуклеоид, и никакими мембранами не окружена. Окрашивание клеток T. magnifica DAPI (это вещество специфически метит только ДНК) показало, что ДНК у этой бактерии находится внутри окруженных мембраной структур. В этих же компартментах концентрируются рибосомы. Пузырьки, содержащие геномную ДНК, разбросаны по всей цитоплазме клетки, включая апикальные почки. Подобные мембранные компартменты, содержащие и геномную ДНК, и рибосомы, ранее не были описаны ни у одной бактерии! Ученые предложили называть эти органеллы пепинами (pepins, от английского слова pips, которым называют мелкие косточки фруктов, например, киви или арбуза) [1].
Стоит уделить внимание и тому, как организована ДНК внутри этих бактерий. В наиболее типичном случае бактериальная клетка содержит единственную копию генома, представленную кольцевой молекулой ДНК. Однако увеличение размеров клетки неизбежно приведет к тому, что молекулы РНК, считанные с одной-единственной копии генома, а также синтезированные на их матрице белки просто физически не смогут достичь удаленных концов клетки. Именно по этой причине все гигантские бактерии полиплоидны и содержат от нескольких десятков до нескольких тысяч (!) копий генома. T. magnifica в этом отношении не стала уникальной, хотя степень ее полиплоидии поражает воображение: на 1 миллиметр нитевидной клетки приходится около 37 тысяч копий генома! Внутри двухсантиметровой клетки находится около 740 тысяч копий генома. Стоит ли говорить, что это абсолютный рекорд по количеству копий генома на клетку среди бактерий. Колоссальная полиплоидия открывает беспрецедентные возможности для геномных перестроек в пределах одной клетки, а именно, всевозможных дупликаций и подобных изменений [1].
Секвенирование генома T. magnifica и сравнение полногеномных последовательностей отдельных клеток показал, что популяции T. magnifica гомогенны на геномном уровне, как обычно и бывает в случае полиплоидных бактерий. Для секвенирования были использованы пять клеток, прикрепленных к одному и тому же листу мангрового кустарника и, следовательно, представляющих одну популяцию. Вывод о гомогенности популяций бактерии был сделан на основании сравнения геномов секвенированных клеток: оказалось, что доля идентичных нуклеотидов в одинаковых позициях составляет целых 99,5%! Сам геном T. magnifica состоит из примерно 12 миллионов пар оснований, что вдвое превышает длину генома второго представителя рода Thiomargarita с секвенированным геномом, Candidatus T. nelsonii. При среднем размере бактериальных геномов в 4,2 миллиона пар оснований геном T. magnifica является одним из крупнейших секвенированных геномов бактерий. В его состав входит 11,788 генов, при том что средний геном бактерии включает около 4 тысяч генов. По размеру генома T. magnifica сопоставима с пекарскими дрожжами Saccharomyces cerevisiae, чей геном включает 12,1 млн пар оснований, а по числу генов эта бактерия обходит даже гриб Aspergillus nidulans, в геноме которого насчитывается порядка 9,5 тысяч генов [1].
Судя по функциональной аннотации генов, T. magnifica является хемоавтотрофным организмом, способным к фиксации углерода и окислению соединений серы, что подтверждается наличием в ее цитоплазме гранул элементарной серы. Примечательно, что, несмотря на внушительный арсенал метаболических путей, T. magnifica практически полностью лишена генов, ответственных за восстановление нитрата и денитрификацию. Таким образом, T. magnifica может использовать нитрат как акцептор электронов в дыхательной цепи. Отсутствие эпибиотических бактерий на поверхности клеток T. magnifica можно объяснить продукцией большого количества вторичных метаболитов, которые, возможно, обладают активностью антибиотиков. В геноме T. magnifica закодированы десятки нерибосомных пептидсинтаз и поликетидсинтаз, которые и отвечают за синтез веществ такого рода [1].
Необычная морфология T. magnifica отразилась и на репертуаре генов, продукты которых участвуют в делении клеток. Оказалось, что T. magnifica утратила львиную долю генов, необходимых для деления бактериальных клеток, однако имеет полный набор генов, связанных с удлинением клеток. Возможно, именно сокращение количества генов, отвечающих за деление, вкупе с активным удлинением клеток лежит в основе формирования невероятно длинных клеток [1].
Ранее мы отмечали, что на концах нитей T. magnifica находится от одной до четырех апикальных почек. Наблюдения за клетками T. magnifica в лаборатории показали, что эти почки время от времени отваливаются и дают начало новым нитевидным клеткам. Таким образом, компактные апикальные почки можно рассматривать как стадию жизненного цикла, на которой возможно распространение T. magnifica в пространстве: зрелые клетки, длинные и тонкие, слишком хрупки, чтобы служить надежной формой распространения. К слову, жизненный цикл с неравным делением и формированием почек встречается и у других бактерий, например, Caulobacter crescentus. Поскольку деление у T. magnifica неравное, апикальные почки получают лишь малую толику копий генома бактерии, которые располагаются внутри пепинов в апикальных почках. Если рассматривать апикальные почки как дочерние клетки, то жизненный цикл T. magnifica напоминает таковой у миксобактерий, формирующих плодовые тела, стрептомицет и даже инфузории Zoothamnium niveum. Авторы работы отмечают, что в этом случае имеет место конвергентная эволюция жизненных циклов у представителей разных доменов [1].
В заключение хотелось бы немного порассуждать о том, почему такие огромные бактерии, как T. magnifica, действительно ломают наши стереотипы о привычных размерах организмов из разных групп. Не секрет, что размер организма ограничивается множеством факторов окружающей среды. Например, почему сейчас не бывает двухметровых стрекоз, которые населяли Землю в каменноугольном периоде? За доставку кислорода к тканям у насекомых отвечает система тонких трубочек — трахей, которые буквально пронизывают тело животного. В карбоне уровень кислорода был выше, чем сейчас, поэтому эффективная доставка кислорода посредством трахей была доступна даже для очень больших насекомых. С современным уровнем кислорода гигантские насекомые бы не выжили: трахеи не смогли бы доставить необходимое количество кислорода ко всем тканям и органам [4]. В случае с прокариотическими организмами ограничения, накладываемые на размер, еще более строгие. У прокариот нет систем активного внутриклеточного транспорта, и молекулы движутся в цитоплазме, полагаясь на пассивную диффузию, которая хорошо подходит для переноса молекул в пределах нескольких микрометров. Очевидно, что в больших клетках диффузия не сможет обеспечивать эффективный транспорт разнообразных молекул во все уголки клетки. Большая клетка требует огромного количества белковых молекул, и, чтобы обеспечить его синтез, необходимо такое количество рибосом, что клетка будет буквально забита ими. Наконец, изменение соотношения площади поверхности клетки к ее объему приводит к тому, что, даже если заполнить всю мембрану молекулами АТФ-синтазы (напомним, АТФ-синтаза — это фермент, который всегда заякорен в мембране), энергии для такой огромной клетки все равно будет не хватать. При этом T. magnifica в три раза превышает максимальный теоретический размер прокариотической клетки, рассчитанный с учетом перечисленных ограничений. Как же ей это удается? Вполне возможно, что усложнение внутриклеточной организации, а именно, наличие многочисленных мембранных органелл, помогает бактерии «обмануть» законы природы. Как знать, может, в скором времени мы узнаем о еще более крупных и странных бактериях и археях, которые полностью сломают наши представления об устройстве прокариотической клетки.
Литература
- Jean-Marie Volland, Silvina Gonzalez-Rizzo, Olivier Gros, Tomáš Tyml, Natalia Ivanova, et. al. A centimeter-long bacterium with DNA compartmentalized in membrane-bound organelles — Cold Spring Harbor Laboratory;
- Гиганты вирусного мира;
- H. N. Schulz, T. Brinkhoff, T. G. Ferdelman, M. Hernández Mariné, A. Teske, B. B. Jørgensen. (1999). Dense Populations of a Giant Sulfur Bacterium in Namibian Shelf Sediments. Science. 284, 493-495;
- R Dudley. (1998). Atmospheric oxygen, giant Paleozoic insects and the evolution of aerial locomotor performance.. Journal of Experimental Biology. 201, 1043-1050.



Комментарии
0Чтобы оставить комментарий, необходимо
войти