Чего вы не знаете о рибосоме...
30 октября 2018
Чего вы не знаете о рибосоме...
- 13174
- 0
- 10
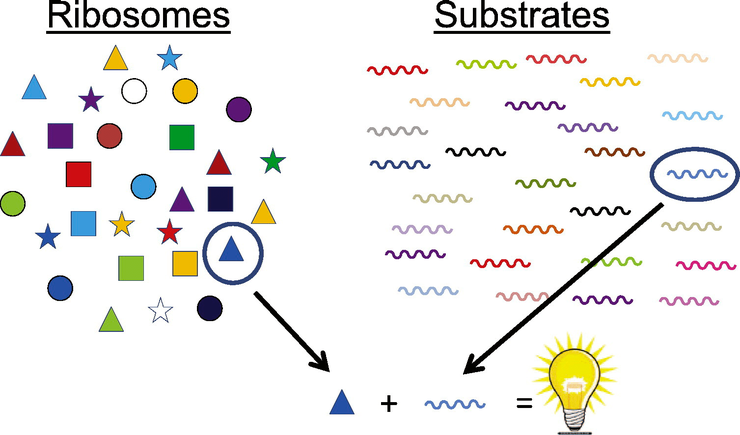
Рибосомы обладают избирательностью
-
Автор
-
Редактор
Статья на конкурс «био/мол/текст»: Еще со школьной скамьи мы знакомы с рибосомой — внутриклеточным органоидом, производящим белки. Роль рибосомы давно перестала быть тайной для человечества. Тут все просто и понятно: используя матричную РНК в качестве «инструкции по сборке»: рибосома последовательно соединяет аминокислоты в длинную белковую цепь. Казалось бы, назначение этого органоида в клетке — разгаданная загадка, и интерес к рибосоме должен вот-вот угаснуть. Но в последние десятилетия в области изучения рибосом совершается колоссальный переворот. И мы можем наблюдать его своими глазами!

Конкурс «био/мол/текст»-2018
Эта работа опубликована в номинации «Свободная тема» конкурса «био/мол/текст»-2018.
Генеральный спонсор конкурса — компания «Диаэм»: крупнейший поставщик оборудования, реагентов и расходных материалов для биологических исследований и производств.
Спонсором приза зрительских симпатий выступил медико-генетический центр Genotek.
«Книжный» спонсор конкурса — «Альпина нон-фикшн»
Белки принимают участие в самых разных клеточных процессах. Например, они могут быть строительными блоками для образования клеточных структур, могут катализировать протекание химических реакций в клетке, осуществлять транспорт молекул из клетки и обратно, а также выполнять некоторые другие важные функции. Понятно, что для нормального роста и развития любой клетке необходимо синтезировать белки. А для их производства, как известно, необходимы рибосомы, ведь именно они отвечают за этот процесс.
Рибосома эукариот состоит из двух субъединиц — большой (ее принято обозначать 60S) и малой (40S). Субъединицы построены из рибосомной РНК (рРНК) и рибосомных белков (РБ). Всего в эукариотической рибосоме находятся около 80 рибосомных белков и 4 типа рРНК .
Об экспериментах ученых над субъединицами рибосом читайте в статье «Биомолекулы» «Две рибосомные субъединицы объединили в функциональный гибрид» [1].
Последовательность, по которой рибосома должна соединить отдельные аминокислоты в белок, записана в молекуле ДНК в виде кода. ДНК состоит из структурных единиц — нуклеотидов. Тройка нуклеотидов образует кодон и кодирует одну аминокислоту . Но как «коду», находящемуся в ДНК, попасть к рибосомам? На помощь приходит молекула-посредник — матричная рибонуклеиновая кислота (мРНК). Она синтезируется по молекуле ДНК, как бы копируя ее последовательность (процесс называется транскрипцией), а затем перемещается к рибосоме. Рибосома в свою очередь осуществляет процесс трансляции (или декодирования), то есть переводит нуклеотидную последовательность в последовательность аминокислотную . Этот процесс отражает центральную догму молекулярной биологии и описывает механизм образования белка [2].
У «Биомолекулы» есть интересная статья-рассуждение о том, как именно мог сложиться этот «кодовый» язык»: «У истоков генетического кода: родственные души» [3]. Интересно, что генетический код может эволюционировать — «Биомолекула» приоткрывала завесу этой тайны в статье «Эволюция генетического кода» [4].
О том, как ученые исследовали это процесс, «Биомолекула» рассказывала в статье «Рибосома за работой» [5].
Еще несколько увлекательных статей «Биомолекулы» о генетическом коде (и не только) для самых любознательных читателей: «Самые нестандартные генетические коды» [6] и «Таинственный код нашего генома» [7].
Количество рибосом в клетке достаточно велико — более 15 тысяч на одну клетку. Одна рибосома синтезирует белок со скоростью 15–20 аминокислот в секунду. При этом ошибается она достаточно редко — одна ошибка случается на 3000 аминокислот. Если принять, что белки в среднем состоят из 400 аминокислотных остатков, то получается, что рибосома совершает всего 1 ошибку на 7 белков! Таким образом, работа рибосомы — высокоточный процесс, в котором задействовано множество дополнительных молекул. Известный американский биохимик Брюс Альбертс даже сравнил рибосому с «молекулярной машиной», подчеркнув этим слаженность и элегантность работы этого сложного молекулярного комплекса [8].
Рибосома была открыта в 1955 году [9], в то самое время, когда ученые ломали голову над тем, как именно генетическая информация перекодируется из ДНК в белки. Интересно, что уже тогда исследователи обратили внимание на неодинаковость рибосом в клетке (Джордж Паладе заметил, что размеры и формы клеточных рибосом немного различаются [10]). Предполагалось, что эта неодинаковость вызвана их различным составом. Фрэнсис Крик, являясь сторонником данного предположения, даже выдвинул гипотезу, суть которой можно выразить фразой «один ген — один белок — одна рибосома» [11]. Он полагал, что существует некий РНК-посредник (в то время о существовании мРНК еще не было достоверно известно), который после синтеза на ДНК встраивается в рибосому, а она, используя «код» этой РНК, производит белок. Предполагалось, что в клетке должны присутствовать тысячи различных рибосом, каждая из которых «сделана под заказ» для производства одного-единственного белка с использованием одного-единственного РНК-посредника. Но через некоторое время эту гипотезу признали ошибочной. Поскольку главные вопросы молекулярной биологии того времени относились к реализации генетической информации, изучение феномена неодинаковости рибосом отошло на второй план.
Новые вехи в истории рибосомы
Новая волна интереса к рибосомам поднялась в начале 90-х годов прошлого столетия. Возможно, толчком к этому послужили две примечательные работы, посвященные исследованию рибосом на разных стадиях жизни эукариотических организмов — малярийного плазмодия и клеточного слизевика.
Малярийный плазмодий вызывает у человека тяжелое заболевание — малярию. Заражение происходит после укуса малярийного комара, в слюне которого находится возбудитель болезни. Отличительной особенностью плазмодия является то, что его жизненный цикл состоит из двух частей (рис. 1). Одна часть протекает в теле комара, другая — в теле человека [12].

Рисунок 1. Жизненный цикл малярийного плазмодия
[12], рисунок с изменениями
После укуса спорозоиты плазмодия — мелкие червеобразные подвижные клетки — попадают в кровоток. По кровотоку они достигают печени, где размножаются бесполым путем, превращаясь в одноядерные клетки, называемые мерозоитами. Голодные мерозоиты атакуют клетки крови — эритроциты — и проникают в них. Питаясь гемоглобином, мерозоиты продолжают бесполое размножение. Через некоторое время они разрушают эритроциты и снова выходят в кровь, где набрасываются на новые эритроциты. Это происходит циклично через каждые 48 часов. При разрушении эритроцитов в кровоток попадают и токсичные продукты жизнедеятельности мерозоитов, так что у человека через равные промежутки времени наблюдаются приступы лихорадки с ознобом и высокой температурой.
На стадии выхода из эритроцитов часть мерозоитов приобретает признаки половых клеток (мужских и женских) — образуются гаметоциты. Когда это происходит, плазмодий вступает во вторую стадию жизненного цикла, которая протекает в теле комара. Если комар выпивает кровь больного малярией человека, то в его организме гаметоциты окончательно созревают и превращаются в гаметы. Именно в теле комара гаметы сливаются, образуя оплодотворенную яйцеклетку. Для дальнейшего развития яйцеклетке необходимо попасть в кишечник комара, где она, проходя несколько стадий, образует спорозоиты. Малярийный плазмодий возвращается в то состояние, когда для дальнейшего развития ему снова необходимо попасть в тело человека. Как только комар кусает свою жертву, спорозоиты попадают в кровь, и цикл повторяется.
Так вот, изучая рибосомы малярийного плазмодия, ученые выяснили, что рРНК малой субъединицы рибосомы кодируется двумя генами [13]. Оказалось, что в клетках плазмодия, живущего в москитах, образуется продукт одного рРНК-гена, а в клетках, попавших в млекопитающее или человека, — продукт другого гена рРНК. Выходило, что, во-первых, состав рибосомы может меняться, а, во-вторых, что эти изменения зависят от состояния внешней среды и от фазы жизненного цикла организма.
Похожие результаты, но уже в отношении других компонентов рибосомы — рибосомных белков — несколькими годами позже получили другие ученые.
Dictyostelium discoideum — слизевик, обитающий в верхних слоях почвы, — является излюбленным модельным организмом молекулярных биологов.
Бóльшую часть жизненного цикла диктиостелиум проводит в виде одноклеточных амёбоподобных форм (миксамёб), питающихся бактериями и дрожжами. При благоприятных условиях (достаточная влажность, нужная температура, наличие питательных веществ) миксамёбы питаются и свободно размножаются делением надвое. Можно сказать, что миксамёбы — индивидуалисты, предпочитающие избегать контактов с сородичами, но когда их численность возрастает и пищевые ресурсы заканчиваются, начинают происходить удивительные вещи. Одна или несколько голодающих клеток принимаются выделять молекулы циклического аденозинмонофосфата (цАМФ) — сигнала «клеточного голода», который одновременно является хемоаттрактантом, привлекающим к источнику сигнала соседних миксамёб. Во время миграции ползущие миксамёбы выделяют и собственное небольшое количество цАМФ, привлекая все больше и больше новых клеток. В результате происходит быстрое объединение огромного количества миксамёб в единый холмообразный агрегат, насчитывающий до нескольких тысяч особей. Так возникает псевдоплазмодий — большое скопление амёбоидных клеток. От обычного плазмодия он отличается тем, что образующие его клетки не сливаются воедино полностью, а, находясь в тесном контакте с соседними клетками, все же сохраняют обособленность и индивидуальность. После агрегации, клетки, находящиеся на вершине «холма», все еще выделяют сигнальные молекулы, и за счет этого продолжают выталкиваться кверху клетками, находящимися у «подножия». Плазмодий вытягивается вверх, а затем заваливается на бок. С этого момента он переходит в стадию «слизня». Мигрируя по направлению к свету и более высокой температуре, слизень достигает поверхности почвы, где в результате перегруппировки и дифференцировки составляющих его клеток формируется плодовое тело. Оно состоит из вытянутого стебелька и находящейся на его вершине споровой коробочки. Споры после созревания рассеиваются ветром, и на новых местах при благоприятных условиях из них выходят миксамёбы, а жизненный цикл слизевика начинается заново (рис. 2) [14].

Рисунок 2. Этапы жизненного цикла слизевика Dictyostelium discoideum
Именно этот необычный жизненный цикл диктиостелиума заинтересовал ученых из Института молекулярной биологии в Нью-Джерси [15]. Они исследовали рибосомы клеток, находящихся на разных жизненных этапах: на стадии одноклеточной миксамёбы (назовем их рибосомы «старыми») и на стадии споры (назовем их рибосомы «новыми»). Из клеток обеих стадий выделили рибосомные белки и количественно сравнили. Результаты были неожиданными. Как оказалось, между стадиями жизненного цикла происходит некое «переключение» состава рибосом, когда «старые» рибосомы утилизируются и на их место приходят «новые», состав которых отличен от первоначального (рис. 3)! В чем состояло отличие? Оказалось, что «новые» рибосомы содержат иные количества некоторых рибосомных белков по сравнению со «старыми». Опять, как и в случае с плазмодием, получалось, что состав рибосом меняется в зависимости от стадии клеточного цикла, но изменениям в данном случае подвергается уже не рРНК, а другой компонент рибосомы — белки.

Рисунок 3. Изменение клеточного состава рибосом слизевика Dictyostelium discoideum в течение жизненного цикла
[12], рисунок с изменениями
Как можно объяснить полученные в ходе этих двух экспериментов данные, фиксирующие изменения в составе рибосом, которые зависят от стадии жизни организма и от условий внешней среды? Знакомясь с научными статьями, посвященными этому вопросу, можно встретить разные рассуждения. Например, такое. Оно опирается на наше знание того, что на разных стадиях жизненного цикла производимый клеткой набор белков примерно схож. Но для каждой стадии существуют и свои характерные белки. По-видимому, для производства таких белков на каждой конкретной стадии необходимы «специальные» рибосомы. При переходе на другую стадию, где требуются иные характерные белки, образуются иные «специальные» рибосомы. Но как рибосома узнает, какие именно белки ей производить? Можно предположить, что для производства «нужных» белков она должна «выбрать» соответствующие им мРНК из всех мРНК, присутствующих в клетке. Как и какие мРНК ей выбрать для трансляции? Опять же можно предположить, что в рибосоме существует некий участок (это могут быть области рРНК, рибосомных белков или те и другие одновременно), взаимодействующий с молекулами мРНК и каким-то образом производящий «селекцию» тех молекул мРНК, которые будут использованы для трансляции. Иными словами, те или иные молекулы, входящие в рибосому, могут напрямую влиять на то, какие мРНК она «выберет» для трансляции и, следовательно, на то, какие белки будут образовываться в клетке [16]. Эти рассуждения приводят к вероятности существования взаимозависимости между набором белков и составом рибосом.
Встречаются и другие рассуждения. Вот еще одно из них. Обновление молекул, включая и те, что входят в рибосому, — это главнейший процесс во время роста и развития клетки. Поэтому можно рассматривать синтез «новых» рибосом как механизм поддержания стабильности этих органелл и их функциональной целостности для обеспечения выживаемости организма [17]. Например, в случае со слизевиком ученые обнаружили, что малая (40S) субъединица рибосом у спор более устойчива к воздействию температуры, чем малая субъединица рибосом миксамёб. Возможно, именно «новые» рибосомные белки обеспечивали эту стабильность. Эти рассуждения могут приводить к мысли о том, что рибосомы способны играть роль в поддержании гомеостаза для повышения выживаемости организма.
Во времена описываемых опытов образование зрелой 80S рибосомы из двух субъединиц являлось загадкой, но третий вариант рассуждений [16] был приближен к современному и опирался на то, что для соединения двух субъединиц в одну зрелую крайне важно взаимодействие рРНК и рибосомных белков. Однако было непонятно, какую именно роль играют рРНК и рибосомные белки в образовании ее функциональных областей. Предполагалось, что рРНК крайне важна для производства белковой цепи, и ее «правильная» структура необходима для протекания разных этапов этого процесса. А вот рибосомные белки нужны именно для создания такой структуры. Они являются каркасом для правильного расположения рРНК в зрелой рибосоме. Таким образом, назначение «переключения» белкового состава рибосом — регулировать пространственную структуру рибосомы, обеспечивая доступ к ее участкам, необходимым для трансляции белков именно на данном этапе жизненного цикла.
Проведенные опыты позволили ввести понятие гетерогенной популяции рибосом — то есть такой популяции, в которой один или несколько структурных компонентов (рибосомные белки или рибосомные РНК) зрелой органеллы подвергались изменениям (качественным или количественным) в течение жизненного цикла клетки.
Возникало множество вопросов. Например, какое из приведенных выше суждений справедливо, какая теория верна? Какую все-таки роль играют рибосомные белки и рРНК в функционировании рибосомы? Влияют ли они на синтез специфических молекул, характерных для клеток определенных стадий и тканей? Каков механизм «переключения» состава рибосомы? Существуют ли гомогенные популяции рибосом, то есть такие, состав которых не меняется? А также возникали многие другие вопросы, ответов на которые в то время не было.
Рибосома — молекулярный фильтр?
В данной статье невозможно описать все эксперименты, проведенные исследователями во время поиска ответов на эти вопросы, хотя многие из них были крайне смелыми и новаторскими и, несомненно, увлекли бы читателя. Но мы ограничимся наиболее значительными, а также опишем ключевые научные идеи, лежащие в основе современного взгляда на рибосому.
В самом начале нового тысячелетия возникла гипотеза рибосомного фильтра [18]. Заслуга авторов этой гипотезы, Винсента Мауро и Геральда Эдельмана, в том, что они, проанализировав и обобщив накопившиеся к тому времени данные, предложили совершенно новый взгляд на рибосому и на принцип ее функционирования. Если раньше рибосома рассматривалась как пассивная структура, с машинной точностью воспроизводящая генетическую информацию, то новый взгляд, предложенный авторами, возводил рибосому в новый ранг, и теперь ей приписывалась неожиданная роль в управлении фундаментальными клеточными процессами. Она приобретала звание «фильтра», который из общего потока клеточных мРНК отсеивает некоторую часть с заданными свойствами и на их основе производит белок. Остальные же мРНК она для трансляции не использует. Авторы предложили и механизм «отсеивания».
Дело в том, что в мРНК есть не только области, кодирующие аминокислотную последовательность белка, но и области, имеющие чисто регуляторное назначение (рис. 4). С их помощью начинать синтезировать белок рибосома может по двум разным механизмам.

Рисунок 4. Белки могут образовываться двумя разными способами. Слева изображен кэп-зависимый механизм начала трансляции. Рибосома садится на кэп, который располагается на одном из концов мРНК, в так называемой нетранслируемой области (НТО), так как она не «переводится» в белок, а является чисто регуляторной. Справа — кэп-независимый механизм. Рибосома садится на IRES, который, благодаря своей сложной пространственной структуре (в рамке), направляет ее к старт-кодону.
рисунок автора статьи
Первый носит название кэп-зависимого, или сканирующего. На одном из концов молекулы мРНК имеется специальная структура — кэп (молекула 7-метилгуанозина), на который перед началом синтеза садится рибосома. Сев, она движется вдоль молекулы мРНК, сканируя ее на предмет старт-кодона, кодирующего первую аминокислоту белка. Найдя старт-кодон, рибосома начинает синтез.
При втором механизме, кэп-независимом, характерном в основном для вирусов, рибосома садится на специальную область, называемую IRES (Internal Ribosomal Entry Site; в переводе с англ. — «сайт внутренней посадки рибосомы»), которая и направляет ее к старт-кодону.
Так вот, гипотеза рибосомного фильтра предполагала, что «фильтрование» происходит за счет специфического взаимодействия рибосомных белков и рРНК с определенными областями мРНК, которые могут запускать синтез белка по схожему с IRES принципу. Такие взаимодействия должны носить конкурирующий характер. Это значит, что мРНК, обладающие одинаковыми или схожими областями, будут соперничать за связывание с рибосомой, и эффективность их трансляции будет различна.
Способность клетки менять состав рибосом также имеет огромное значение для работы «фильтра». Добавление или утрата некоторых компонентов может приводить к пространственной недоступности областей рибосомы, взаимодействующих с мРНК, а значит, будет блокировать и синтез некоторых белков. Именно так, по мнению авторов гипотезы, могла меняться специфичность «фильтра».
Гипотеза рибосомного фильтра породила другую гипотезу — о специализации рибосом [19]. Специализация — следствие гетерогенности, то есть избирательность рибосом в отношении мРНК, которая приводит к синтезу только каких-то определенных белков, закодированных в «избранных» рибосомой молекулах мРНК. К сожалению, ввиду отсутствия приборов и методов, с помощью которых можно было бы экспериментально проверить описанные идеи, эти гипотезы так и оставались гипотезами некоторое время.
Гипотезы приобретают подтверждение
Несомненно, двадцать первый век — век высоких технологий, позволяющих человеку совершать то, что раньше казалось выдумкой ученых-фантастов. И молекулярные биологи активно пользуются результатами научно-технического прогресса в своих научных изысканиях. Благодаря интенсивному развитию техники, несколько лет назад сразу обе гипотезы нашли свое экспериментальное подтверждение [20]. С помощью самых современных технологий установили, что рибосомы модельного объекта — стволовой клетки эмбриона мыши, не обладающей таким сложным жизненным циклом, как клетки диктиостелиума или малярийного плазмодия, также гетерогенны, то есть отличаются друг от друга составом входящих в них белков. Получается, что и клетки млекопитающих содержат неодинаковые по составу рибосомы!
Дальше — больше. Оказалось, что в одной и той же клетке присутствует не одна гетерогенная популяция рибосом, как полагали ранее, а сразу несколько таких популяций! Они не сменяют друг друга, а сосуществуют одновременно. Было экспериментально доказано, что каждая из популяций преимущественно осуществляет трансляцию только определенного набора мРНК. Это означает, что рибосомы не универсальны, то есть не способны транслировать абсолютно любую молекулу мРНК, как считалось ранее. Отдельные группы рибосом взаимодействуют только с некоторыми мРНК и, следовательно, «отвечают» за синтез лишь части клеточных белков. Было доказано, что избирательность рибосом вызвана именно присутствием определенных рибосомных белков, которые, предположительно, связываются с некими регуляторными областями в мРНК. Но точный механизм этого процесса требует дальнейших исследований.
Итак, получается, что функции рибосомных белков более обширны, чем мы были склонны думать ранее! Благодаря изменениям состава рибосомы клетка может регулировать, какие гены будут «превращены» в белки, а какие нет, в соответствии с ее нуждами.
Что же дальше?
Пока рано говорить о том, куда приведут нас исследования гетерогенности рибосом. Несомненно, они позволят человечеству лучше понимать механизмы многих заболеваний, в том числе онкологических.
Например, уже выявлены заболевания, именуемые рибосомопатиями, причиной которых являются генетические мутации компонентов рибосом [21].
Анемия Даймонда—Блэкфана — типичный пример рибосомопатии, выявляется в течение первого года жизни ребенка. Это заболевание названо в честь врачей, впервые его описавших и классифицировавших в 30-е годы двадцатого века. Оно относится к числу редких, встречается в пяти случаях из миллиона, с одинаковой вероятностью и у мальчиков, и у девочек. Первичным симптомом болезни является повышенная бледность кожи ребенка, описаны случаи рождения детей с «заячьей губой». К возрасту шести месяцев могут появиться такие симптомы как увеличение размеров печени и селезенки, нарушения работы сердечно-сосудистой системы. Также происходит процесс торможения выработки эритроцитов в костном мозге, из-за чего наблюдаются бледность, вялость, быстрая утомляемость. При отсутствии лечения заболевание приводит к тяжелой сердечной недостаточности и смерти. Как оказалось, причиной, вызывающей анемию Даймона—Блэкфана, являются мутации в генах, кодирующих некоторые рибосомные белки, что приводит к неправильной работе рибосом, снижению их количества в клетках и, как следствие, негативным последствиям.
В заключение
История рибосомы, начавшаяся более шестидесяти лет назад, — яркий пример процесса научного поиска, который привел к изменению нашего взгляда на работу и функции рибосомы. Сейчас мы не можем сказать, чем закончится эта история, к каким результатам и открытиям она приведет.
В науке не бывает незначительных результатов. Мы, обыватели, привыкли, что ее достижения влияют на нашу жизнь, а, следовательно, должны быть масштабными и иметь непосредственное практическое применение. Например, открытие пенициллина — первого антибиотика — позволило спасти миллионы жизней и дало виток новому направлению исследований, до сих пор не утратившему актуальности. Масштаб и польза налицо. Однако нам следует помнить, что в области познания не бывает «больших» и «малых» достижений. Любой научный результат — это всегда движение вперед, это всегда большая победа, созданная огромным кропотливым трудом целой плеяды ученых. Так же и исследования рибосомы. Возможно, они станут еще одним штрихом, добавленным учеными к нашей картине мира, а возможно, приведут к колоссальному прорыву, например, в лечении онкологических заболеваний.
Литература
- Две рибосомные субъединицы объединили в функциональный гибрид;
- Грайфер Д.М, Карпова Г.Г., Малыгин А.А. (2006). Рибосома — минифабрика по производству белков. «Наука из первых рук»;
- У истоков генетического кода: родственные души;
- Эволюция генетического кода;
- Рибосома за работой;
- Самые нестандартные генетические коды;
- Таинственный код нашего генома;
- Bruce Alberts. (1998). The Cell as a Collection of Protein Machines: Preparing the Next Generation of Molecular Biologists. Cell. 92, 291-294;
- G. E. Palade. (1955). A SMALL PARTICULATE COMPONENT OF THE CYTOPLASM. The Journal of Cell Biology. 1, 59-68;
- Palade G.E. (1958). Microsomes and ribonucleoprotein particles. Microsomal Particles and Protein Synthesis, First Symposium of the Biophysical Society. 36–61;
- Crick F.H.C. (1958). On protein synthesis. Symp. Soc. Exp. Biol. 12, 138–163;
- Shifeng Xue, Maria Barna. (2012). Specialized ribosomes: a new frontier in gene regulation and organismal biology. Nat Rev Mol Cell Biol. 13, 355-369;
- J. Gunderson, M. Sogin, G Wollett, M Hollingdale, V. de la Cruz, et. al.. (1987). Structurally distinct, stage-specific ribosomes occur in Plasmodium. Science. 238, 933-937;
- Bernard Girau, César Torres-Huitzil, Nikolaos Vlassopoulos, José Hugo Barrón-Zambrano. (2009). Reaction Diffusion and Chemotaxis for Decentralized Gathering on FPGAs. International Journal of Reconfigurable Computing. 2009, 1-15;
- Ramagopal S. and Ennis H.L. (1982). Ribosomal protein synthesis during spore germination and vegetative growth in Dictyostelium discoideum. J. Biol. Chem. 257,1025–1031;
- S. Ramagopal. (1992). Are eukaryotic ribosomes heterogeneous? Affirmations on the horizon. Biochem. Cell Biol.. 70, 269-272;
- Subbanaidu Ramagopal, Herbert L. Ennis. (1984). Decay and synthesis of ribosomal proteins during Dictyostelium discoideum development. Molec. Gen. Genet.. 194, 466-470;
- V. P. Mauro, G. M. Edelman. (2002). The ribosome filter hypothesis. Proceedings of the National Academy of Sciences. 99, 12031-12036;
- Jonathan D. Dinman. (2016). Pathways to Specialized Ribosomes: The Brussels Lecture. Journal of Molecular Biology. 428, 2186-2194;
- Zhen Shi, Kotaro Fujii, Kyle M. Kovary, Naomi R. Genuth, Hannes L. Röst, et. al.. (2017). Heterogeneous Ribosomes Preferentially Translate Distinct Subpools of mRNAs Genome-wide. Molecular Cell. 67, 71-83.e7;
- A. Narla, B. L. Ebert. (2010). Ribosomopathies: human disorders of ribosome dysfunction. Blood. 115, 3196-3205.



Комментарии
0Чтобы оставить комментарий, необходимо
войти