У истоков генетического кода: родственные души
28 июля 2014
У истоков генетического кода: родственные души
- 6516
- 0
- 9
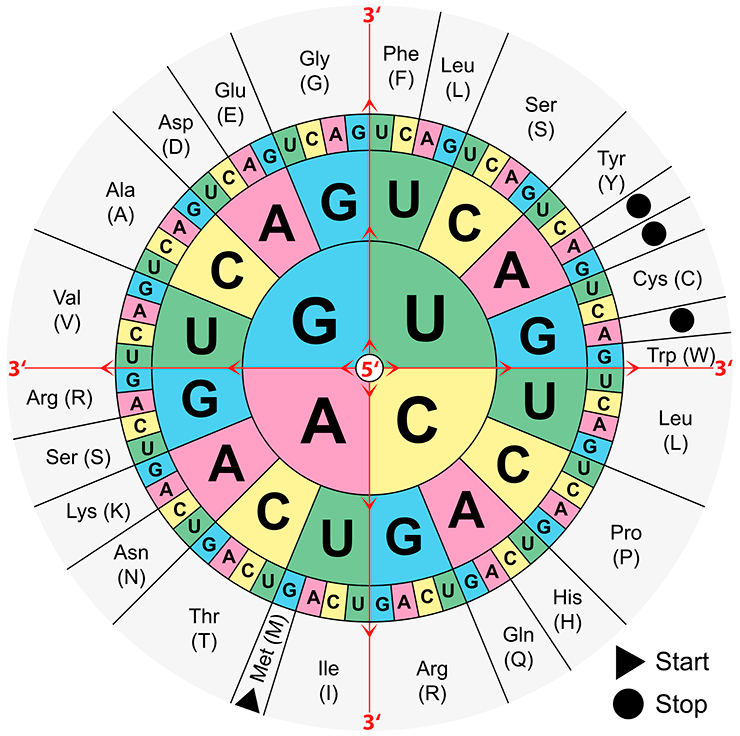
Таблица генетического кода
-
Автор
-
Редакторы
Статья на конкурс «био/мол/текст»: Границы между науками — штука эфемерная. Биология прорастает из химии; физика неразрывно связана с математикой; палеонтология, геология, география, история в тесном сотрудничестве описывают события прежних веков. Огромные массивы биологических данных, полученных с помощью новейших методик исследования, обрабатываются с помощью биоинформатики. И даже такие непохожие науки, как молекулярная биология и лингвистика тоже имеют точки соприкосновения. Не верите? Ну прочитайте статью.

Конкурс «био/мол/текст»-2014
Эта статья представлена на конкурс научно-популярных работ «био/мол/текст»-2014 в номинации «Биоинформатика и молекулярная эволюция».

Главный спонсор конкурса — дальновидная компания Генотек.
Конкурс поддержан ОАО «РВК».
Спонсором номинации «Биоинформатика» является Институт биоинформатики.
Спонсором приза зрительских симпатий выступила фирма Helicon.
Свой приз также вручает Фонд поддержки передовых биотехнологий.
Ну, предположим, мы с вами условимся, что слово «лошадь»
станет для нас означать учебник по грамматике,
тогда мы будем иметь право сказать:
«Откройте вашу лошадь на двадцатой странице» или
«Вы принесли сегодня на занятия свою лошадь?»
И оба прекрасно друг друга поймем, как вы считаете?
Джон Барт. Конец пути
В разных языках одни понятия называются по-разному звучащими словами. По-английски «вода» будет water («вотер»), по-албански — ujë («уё»), по-китайски — 水 («ше»), а по-валлийски — dŵr («дюр»). Наоборот, одинаково звучащее слово в разных языках может означать противоположные вещи: по-русски «яма» — это яма, а по-японски — гора.
Значит, возникновение слова, появление связи между понятием и определяющим его сочетанием звуков или букв — процесс относительно случайный; можно представить, что в русском языке понятие «вода» обозначается словом «соль», «камень», «пиво» или (с большой натяжкой) «аэрофотосъемка».
А теперь посмотрим на генетический код (см. заглавный рисунок). Это тоже язык, только особенный: его буквы —химические соединения, азотистые основания. Этих букв-оснований четыре — гуанин (G), цитозин (C), аденин (A) и урацил (U). Все слова (они называются кодоны) в этом языке трехбуквенные; из четырех букв получается 64 трехбуквенных слова. Эти 64 слова кодируют 21 «понятие»: 20 аминокислот и стоп-кодон. (Очень редко помимо 20 классических аминокислот генетический код кодирует еще две «дополнительных», так что можно сказать, что «понятий» не 21, а 23; но это неважно для нашей истории.)

Рисунок 1. Химическая структура «букв» (нуклеотидов; сверху) и «понятий» (аминокислот; снизу) генетического кода. Помимо двадцати «классических» аминокислот генетический код крайне редко кодирует еще две, одна из которых (селеноцистеин) показана на этом рисунке.
«Википедия»: Nucleobase, Amino acids
И тут возникает заковыристый лингвобиологический вопрос. Является ли генетический код таким же произвольным, случайно образовавшимся языком, как языки, на которых говорят люди? Можно ли представить, что кодоны в генетическом коде кодируют другие аминокислоты, а аминокислоты кодируются другими кодонами? Может ли кодон UUU кодировать не фенилаланин (как он это сейчас делает), а глицин? А кодон AGG — тирозин? А кодон CUC — пролин? Иными словами, случайно ли подбирались кодоны, обозначающие аминокислоты, — или в этом был какой-то смысл?
В последующем тексте я попытаюсь ответить на этот вопрос, но вначале покаюсь. Обсуждение исчезающе далекого прошлого — такое же захватывающее и бесполезное занятие, как обсуждение исчезающе далекого будущего. Ни доказать, ни опровергнуть тут ничего нельзя, и то, что описано в этой статье, — всего лишь гипотетический вариант развития событий. И все-таки обсуждение данных вопросов чрезвычайно расширяет кругозор и тренирует мыслительные способности — так что можно расценивать рассуждения на эту тему как экзаптацию, служащую общему развитию человечества. Эта статья во многом основана на лекции заведующего лабораторией компьютерной биофизики Венского университета Бояна Жагровича [1] во время научной школы-конференции «Современная биология & Биотехнологии будущего», посвященной острым вопросам и актуальным проблемам фундаментальной и прикладной биологии, а также на книге Евгения Кунина «Логика случая» [2] (особенно на главе о происхождении жизни).
Once upon a time...
Представим себе мир на заре возникновения жизни. По самой популярной из существующих сейчас теорий это был РНК-мир [3]: РНК были самыми продвинутыми из существовавших тогда биомолекул. Эти примитивные и неуклюжие «первобытные» РНК тогда занимались и хранением информации (чем сейчас занимается в основном ДНК), и катализом биохимических реакций (чем сейчас занимаются в основном белки). В результате РНК способны были самовоспроизводиться: катализировать на основе одной цепочки РНК создание других цепочек, идентичных исходной. Может быть, еще нельзя было назвать эти молекулы живыми, но уже можно было назвать их бессмертными.
Судя по всему, жизнь зародилась в пористой породе, образующей сеть ячеек, через стенки которых затруднена диффузия [4]. В этой породе находился первичный бульон — водный раствор различных веществ, в том числе этих самых коротких цепочек РНК и отдельных аминокислот. Из-за того, что многим молекулам нелегко было пробраться сквозь стенки ячеек, состав ячеек был разным: в одних по каким-то причинам накапливались одни молекулы, а в других — другие. Такие ячейки можно назвать «протоклетками», потому что их стенки выполняли ту же функцию, которую выполняет мембрана у современных клеток: отделение Мира Внутри от Мира Снаружи.
Разрозненные молекулы, в том числе, РНК и аминокислоты, плававшие в первичном бульоне, могли общаться только одним способом — с помощью физико-химических взаимодействий. Растворимость в воде, электрический заряд, пространственные характеристики, некоторые другие свойства — все это заставляет одни молекулы слипаться друг с другом в растворе, а другие — отплывать друг от друга как можно дальше.
И вот представим себе такую романтическую историю. Некий кодон (сочетание из трех нуклеотидов) и некая аминокислота — это «родственные души». Благодаря своим физико-химическим свойствам они крепко слипаются, встретившись в растворе.
Представим себе, что в некой цепочке РНК есть этот кодон. Есть вероятность, что в той неорганической ячейке, где плавает эта РНК, случайно окажется «родная» для кодона аминокислота. И есть вероятность, что эта аминокислота налипнет на свой «родной» кодон. И есть вероятность, что от этого жизнь той РНК существенно облегчится — она станет стабильнее, или будет лучше работать, или приобретет какие-то новые выгодные свойства. В результате такая РНК станет более приспособленной, и отбор (в тогдашней форме этого процесса) будет ей благоволить.
Это объединение двух невидимых глазу молекул, произошедшее в капле воды в крохотной дырочке пористой породы где-то в океане миллиарды лет назад — гигантский прорыв в истории нашей планеты. С этого момента возникает «дружба» между РНК и белками, краеугольный камень жизни на Земле. С этой дружбы началась биологическая эволюция, благодаря которой существуем и мы с вами.
Допустим, пары «родственных душ» существовали и для других кодонов и аминокислот. Тогда могла сложиться ситуация, когда напротив двух расположенных рядом кодонов РНК встанут две «родные» для этих кодонов аминокислоты. А отсюда — один (правда, довольно трудный) шаг до того, чтобы эти аминокислоты соединились, образовав цепочку из двух звеньев. И если связывание между кодоном и аминокислотой достаточно стабильно, то такая ситуация будет повторяться снова и снова на других молекулах РНК. То есть, напротив одних и тех же сочетаний нуклеотидов будут выстраиваться одни и те же аминокислоты, которые, при счастливом стечении обстоятельств, объединятся в цепочки. Вот так, буквально из ничего, возникает «романтическая связь» между кодонами и аминокислотами — генетический код.
Доказательства
Эмпирическим путем было получено несколько доказательств этого предположения. Еще в 1966 году Карл Вёзе [5] показал, что аминокислоты, кодируемые пиримидиновыми кодонами, имеют повышенное сродство к аналогам пиримидина в растворе [6]. А позже в работах лаборатории Бояна Жагровича было обнаружено, что аминокислоты, кодируемые пуриновыми основаниями, имеют повышенное сродство к пуриновому основанию гуанину (но, почему-то, не к аденину) [7], [8]. Иными словами, имеет значение не столько конкретный нуклеотид, сколько его «половая принадлежность» — пуриновость или пиримидиновость (рис. 2).

Рисунок 2. Зависимость между сродством аминокислоты к аналогам пиримидина в растворе (эмпирическая характеристика под названием polar requirement; подробней о ней рассказано в [9]) и «пиримидиновостью» (дóлей пиримидиновых нуклеотидов) ее кодона. Хотя зависимость не «железная» и исключения налицо, но закономерность явно прослеживается.
Конечно, в таком примитивном виде, без «поддерживающей конструкции» в виде аппарата белкового синтеза, код будет еще очень неточным: легко можно представить себе ситуацию, когда две сходные по свойствам аминокислоты прилипают к одному и тому же кодону. Однако возможно, что в те незапамятные времена еще не требовалась филигранная точность белкового синтеза, и «супружеская измена» (замена одной аминокислоты на другую, обладающую похожими свойствами) несильно меняла свойства тогдашних простейших белковых цепочек.
Шероховатые места
Эту красивую гипотезу омрачает одно обстоятельство. Нить мРНК куда длиннее белковой нити, которую она кодирует. Непонятно, как же куцей аминокислотной цепочке удавалось правильно встать напротив длинной цепочки нуклеотидов. Возможны несколько объяснений.
Во-первых, крайне вероятно, что давным-давно генетический код был не триплетным, а диплетным, и третий нуклеотид в нем появился тогда, когда выросло количество используемых в белках аминокислот (или стала важнее специфичность этих аминокислот). Это предположение подтверждается тем, что часто аминокислота кодируется несколькими альтернативными кодонами, у которых первые два нуклеотида одинаковы, а отличается только последний. Кроме того, когда тРНК распознает свой кодон, самое важное значение имеют первые два нуклеотида, а третий — это уже просто дополнительный «бантик» (см. об этом tRNA wobble). Если предположение о диплетности верно, то соотношение длин нитей мРНК и белка уменьшается, и становится легче представить, как они друг напротив друга устанавливаются.
Во-вторых, цепочка — это сильно сказано. Видимо, во время становления генетического кода речь шла о стабильном присоединении к нужному месту всего одной-двух аминокислот. Удлинение же белковых цепочек шло параллельно с возникновением аппарата белкового синтеза, в том числе рибосомы [10], и проблема несоответствия длин цепочек РНК и белка потеряла свою актуальность.
Дела давно минувших дней
А теперь — небольшое лирическое отступленье о преданьях старины глубокой, порядке возникновения нуклеотидов и некоторых аспектах «дружбы» между РНК и белками.

Рисунок 3. Первичные аминокислоты (выбраны на основе эксперимента Миллера—Юри, показаны красным) имеют особенно высокое сродство к гуанину и почти не имеют сродства к аденину. Остальные, вторичные, аминокислоты (показаны зеленым) имеют куда меньшее сродство к гуанину и ярко выраженное «антисродство» к аденину. Для цитозина и урацила картина напоминает таковую для гуанина, но не столь выражена. Еще приведены данные для всех аминокислот вообще, без разделения на первичные и вторичные (показаны черным), а также общие данные для пуриновых (PUR) и пиримидиновых (PYR) нуклеотидов. По вертикальной оси — коэффициент корреляции между количеством нуклеотида в кодоне и сродством к этому нуклеотиду соответствующей аминокислоты. Исторически сложилось, что в данном случае отрицательное значение коэффициента свидетельствует о высоком сродстве аминокислоты к нуклеотиду и наоборот.
Все аминокислоты можно разделить на первичные и вторичные. Первичные, или эволюционно древние, можно получить в абиотических условиях, они не требуют сложного синтеза, возможного только в живых системах. Вторичные, или эволюционно юные, без этого синтеза получить практически невозможно. Понятно, что в «первобытных» белках могли использоваться только первичные аминокислоты — потому что вторичным неоткуда было взяться, не было ферментов для их создания. И вот оказывается, что первичные аминокислоты, во-первых, чаще имеют в своих кодонах гуанин и цитозин, а во-вторых — если их имеют, то «гуаниновые» показывают большое (больше, чем аминокислоты в среднем) сродство к гуанину, а «цитозиновые» — к цитозину (рис. 3). Возможно, это говорит о том, что пара гуанин—цитозин эволюционно более древняя, чем аденин—урацил; «доисторические» РНК состояли преимущественно из них и кодировали только простые, первичные аминокислоты. (Гуанин и цитозин при спаривании образуют три водородные связи, а аденин и урацил — две; то есть, ГЦ — это более стабильная пара; видимо, «любовь» древних РНК к паре ГЦ связана именно с этим.) Затем появились пути для синтеза вторичных аминокислот, а одновременно помимо двух «первичных» оснований — гуанина и цитозина — в РНК стали чаще появляться «вторичные» — аденин и урацил; в результате именно «вторичные» основания стали кодировать вторичные аминокислоты.
Есть, кстати, интересная зависимость, касающаяся аденина. Показано, что аминокислоты, в кодонах которых много этого нуклеотида, не просто не имеют к своим кодонам сродства, но наоборот, имеют «антисродство»: отталкиваются от аденинов в растворе. Это может говорить о том, что к тому моменту, как в РНК появилось большое количество аденинов (кодирующих сложные вторичные аминокислоты), физико-химические взаимодействия уже потеряли свое решающее значение для трансляции. Либо, возможно, такие аминокислоты показывали сродство не к кодону своему, а к антикодону.
Если исходить из того, что со временем физико-химические взаимодействия теряли свое значение для стабилизации генетического кода, то можно выстроить нуклеотиды в порядке их появления в РНК. В этом случае самым древним нуклеотидом должен быть гуанин — ведь «его» аминокислоты чувствуют к нему особую близость. Косвенным подтверждением этому может служить то, что глицин — самая примитивная (= самая древняя) из существующих аминокислот — кодируется сочетанием из двух гуанинов и еще одного (любого) нуклеотида.
Эхо древнего мира
А теперь посмотрим на вопрос с другой стороны. С тех пор, как за счет физико-химических взаимодействий возник и оптимизировался генетический код, утекло много воды и сменился не один додекальон поколений клеток. Имеют ли эти взаимодействия значение в жизни современной клетки — или их давно уже «загородили» более сильные и «умные» процессы? Звучит ли в современном мире эхо мира древнего?
Может быть, и звучит. Вот всего несколько ситуаций, в которых могут иметь значение прямые физико-химические взаимодействия между белком и РНК.
Во-первых, саморегуляция синтеза белка. Возможно, что связывание едва синтезированной белковой цепочки с породившей ее мРНК предотвращает дальнейшее связывание этой мРНК с рибосомой — а соответственно, и дальнейший синтез такой же белковой цепочки на основе этой мРНК. Получается отрицательная обратная связь. Синтез белка регулируется автоматически — само наличие продукта выключает производство этого продукта. (Но, разумеется, этот гипотетический механизм отнюдь не отменяет большого количества хорошо доказанных механизмов регулировки белкового синтеза.)
Во-вторых, вирусы. В ком (или в чём?) еще белок так тесно соседствует с нуклеиновой кислотой, от которой берет начало? Возможно, белки налипают на нужные участки РНК в РНК-содержащих вирусах как минимум частично за счет вышеописанных физико-химических взаимодействий.
И, наконец, нуклеопротеины — слипшиеся кусочки нуклеиновой кислоты и белка. К ним относятся, например, бурно изучающиеся сейчас P-тельца (см. P-bodies) — облепленные белками нити мРНК, в которых эта мРНК зачастую подвергается деградации. Возможно, что белки облепляют мРНК в том числе и за счет тех древних, прямых физико-химических взаимодействий. Это особенно интересно потому, что слепляться таким способом будут преимущественно неструктурированные белки и РНК (потому что у структурированных «родственные», слипающиеся области как правило спрятаны внутри молекулы). А именно неструктурированные биомолекулы прежде всего и должны подвергнуться уничтожению.
Итог
Итак, каков же ответ на поставленный в начале статьи вопрос? Случайно или неслучайно подбирались кодоны в генетическом коде?
Ответить можно только с оговорками. Общая закономерность проглядывается: между собой соединяются «родственные души». Чем «пиримидиновей» кодон, который кодирует аминокислоту, тем большее сродство эта аминокислота имеет к пиримидинам, чем «пуриновей» — тем охотней аминокислота слипается с пуринами. Однако частности, то, какую именно из «родственных» аминокислот будет кодировать данный кодон, видимо, определялись случайным путем. Не может кодон UUU кодировать глицин — потому что глицин с ним слипаться не будет. Зато он может кодировать что-нибудь «близкое по духу» — лейцин или изолейцин, например.
Мысли в тему
Во всей этой истории есть несколько философских моментов, на которые хочется обратить внимание.
Во-первых, связь между лингвистикой и биологией. При изучении языка можно сделать выводы о происхождении слова, его древности, встречаемости в разные эпохи, изменении значения со временем, не прибегая ни к каким источникам информации, кроме самого языка. Сам язык несет в себе то, что нужно для его изучения. Та же история и с генетическим кодом. И возможно, в исследованиях генетического кода могут пригодиться методы, ныне используемые в лингвистике.
Но, как мы только что выяснили, генетический код, в отличие от человеческого языка, — штука отнюдь не произвольная. Он стал таким, какой есть, не на ровном месте; сами «буквы», в нем используемые (нуклеотиды), своими физико-химическими свойствами неразрывно связаны с «понятиями», которые они определяют (аминокислотами).
И тогда, во-вторых, всплывает еще один философский вопрос: является ли генетический код цифровым или аналоговым?
С одной-то стороны, он, конечно, цифровой — ведь главную информацию несут именно сочетания нуклеотидов, кодоны. Исключительно от последовательности кодонов зависит, какой белок получится на основе РНК. Прочесть эту информацию в клетке просто так невозможно: для этого нужно протянуть всю нить РНК через рибосому и сделать на ее основе белок. Точно так же, как нельзя узнать о том, что происходит в книге, не прочитав ее страницу за страницей.
Но, помимо цифровой, код несет и аналоговую информацию. И чтобы получить эту информацию, клетке никакая рибосома не нужна — информация определяется сама, на основе физико-химических взаимодействий, в которые вступает молекула РНК. Продолжая аналогию — хотя нельзя узнать, что произойдет в книге, не прочтя ее, но сам вид, размер, обложка книги тоже несут какую-то информацию и могут дать подсказки о ее содержимом.
И может быть, описанные в этой задаче закономерности — это только вершина айсберга. Тогда нам предстоит еще многое узнать об аналоговой информации, которую несет генетический код.
Видео. Лекция Бояна Жагровича (Bojan Zagrovic) Computational modeling of biomolecules: goals, achievements and outstanding challenges, прочитанная на Зимней школе «Современная биология & Биотехнологии будущего» (Звенигород, 2014 г.)
Исходно эта статья была опубликована в виде еженедельной задачи по биологии на «Элементах» [11].
Литература
- Zagrovic B. (2014). Computational modeling of biomolecules: goals, achievements and outstanding challenges. YouTube;
- Кунин Е. Логика случая. О природе и происхождении биологической эволюции. М.: «Центрполиграф», 2014. — 528 с.;
- РНК у истоков жизни?;
- К вопросу о происхождении жизни;
- Карл Вёзе (1928–2012);
- C. R. Woese, D. H. Dugre, W. C. Saxinger, S. A. Dugre. (1966). The molecular basis for the genetic code.. Proceedings of the National Academy of Sciences. 55, 966-974;
- Mario Hlevnjak, Anton A. Polyansky, Bojan Zagrovic. (2012). Sequence signatures of direct complementarity between mRNAs and cognate proteins on multiple levels. Nucleic Acids Research. 40, 8874-8882;
- Anton A Polyansky, Mario Hlevnjak, Bojan Zagrovic. (2013). Proteome-wide analysis reveals clues of complementary interactions between mRNAs and their cognate proteins as the physicochemical foundation of the genetic code. RNA Biology. 10, 1248-1254;
- Damien C. Mathew, Zaida Luthey-Schulten. (2008). On the Physical Basis of the Amino Acid Polar Requirement. J Mol Evol. 66, 519-528;
- Элементы: «Тайна происхождения рибосом разгадана?»;
- Элементы: «Родственные души».



Комментарии
0Чтобы оставить комментарий, необходимо
войти