Геном человека: полезная книга, или глянцевый журнал?
11 апреля 2007
Геном человека: полезная книга, или глянцевый журнал?
- 5542
- 1
- 7
-
Автор
-
Редакторы
Темы
Изучение человеческого генома имеет одну конечную цель — оно затевается исключительно ради того, чтобы, взглянув на последовательность ДНК конкретного индивида, можно было бы получить о нем максимум информации. О том, какими болезнями он может заболеть, какие способности в себе развить, и какие опасности его могут поджидать при выборе того или иного жизненного пути. История изучения этого вопроса довольно продолжительна, однако заветная цель приближается к нам далеко не так быстро, как хотелось бы.
История вопроса
Особенности психики человека таковы, что он склонен переоценивать собственные достижения. Эта закономерность очень четко прослеживается в развитии научной мысли. Ученым зачастую кажется, что стоит совершить еще небольшой рывок, как истина откроется во всей красе, а будущим поколениям естествоиспытателей останется лишь стряхивать пыль с приборов. Однако в подавляющем большинстве случаев получается так, что новое открытие порождает больше вопросов, чем дает ответов.
В начале прошлого столетия казалось, что до выяснения природы наследственности рукой подать, ведь были заново открыты законы Менделя, сформулирована Хромосомная теория наследственности Моргана. Согласно представлениям того времени, наследственные факторы — гены — являлись белковыми молекулами, последовательно соединенными между собой в хромосомах. Казалось, что вот-вот эти белки будут выделены из хромосом и все встанет на свои места. При этом, естественно, ни у кого и в мыслях не было, что в генетическом материале организма могут присутствовать элементы, напрямую не определяющие каких-либо его свойств. Была уверенность в том, что каждый ген-белок отвечает за определенную функцию. Однако все оказалось куда сложнее.
Во-первых, к середине 40-х годов XX века, благодаря опытам Эйвери, Маклеода и Маккарти становится понятно, что функции хранения и передачи наследственной информации могут выполнять вовсе не белки. Внимание ученых начинает концентрироваться на изучении ДНК — полимерной молекулы, состоящей из дезоксирибонуклеотидов. К тому моменту было хорошо известно, что ДНК входит в состав хромосом, однако полагали, что эта молекула выполняет структурные функции, являясь своего рода хромосомным каркасом. Окончательно обосновали ключевую роль ДНК в наследственности Альфред Херши и Марта Чейз, только в начале 50-х годов показав, что бактериофаги способны размножаться без собственных белков — в инфицируемой ими бактериальной клетке оказывается и реплицируется только молекула ДНК.
Структуру ДНК впервые описывают Джеймс Уотсон и Френсис Крик в своей работе 1953 года [1]. В последующие 20 лет накапливаются знания о природе генетического кода (М. Ниренберг и Дж. Маттеи), работе генов и регуляторных элементов (Ф. Жакоб и Ж. Моно), тонкой структуре гена (С. Бензер), об укладке ДНК на нуклеосомах (А. Корнберг). Также становится понятно, что в геномах организмов содержатся не только уникальные последовательности структурных генов — в них присутствует огромное количество часто повторяющихся и вовсе не кодирующих белки последовательностей.
Своя и чужая ДНК
В середине прошлого века тезис о том, что генетический материал организма содержит исключительно структуры, необходимые для формирования фенотипических признаков, было странно подвергать сомнению. Любую особенность организма пытались объяснить с позиций целесообразности, и поэтому считалось, что лишних и нефункциональных структур быть просто не должно.
Опыты, подтверждающие наличие в геноме «лишнего» материала, довольно любопытны. Первые указания на такую особенность были получены при изучении кинетики реассоциации геномной ДНК. Дело в том, что последовательности ДНК с разной скоростью восстанавливаются после денатурации. Если последовательность содержит много повторов, она восстанавливается быстрее, а если последовательность ДНК уникальна — времени на ее ренатурацию требуется больше. В ходе проведения экспериментов по реассоциации геномных ДНК эукариот было выяснено, что очень большая часть генома приходится на разного рода быстро ренатурирующие повторы, сателлиты и прочую «бесполезную» ДНК (кстати, термин “junk DNA” был введен еще Френсисом Криком). Еще сравнительно недавно бытовало мнение, что такого рода «мусорные последовательности» необходимы для того, чтобы защищать полезную ДНК от мутаций, как бы вызывая огонь на себя. В самом деле, если мутациям подвергается некодирующий участок ДНК, то это, скорее всего, не отразится негативно на фенотипе особи. Однако, как выяснилось впоследствии, роль такой ДНК в геноме нельзя ограничить только этим. Дальнейшее изучение вопроса показало, что повторяющаяся и некодирующая фракция ДНК в геноме чрезвычайно разнообразна по своей структуре. Помимо повторов, выполняющих чисто технические функции связывания с белками ядерного матрикса или компонентами центромеры, обнаруживаются и такие участки генома, которые напрямую не влияют на морфологию организма и на выполнение им тех или иных функций. Исследование того, откуда эти участки взялись и почему они есть во всех организмах, представляло интерес.

Рисунок 1. Барбара МакКлинток.
Примерно одновременно с работами по изучению роли ДНК в наследственности подвергается первой критике хромосомная теория Моргана в хрестоматийном ее понимании. Это связано с тем, что Барбара МакКлинток обнаруживает генетические элементы, которые, по ее мнению, способны менять свою локализацию на хромосоме [2]. Эти революционные исследования поначалу не находят понимания, поскольку противоречат принятому тогда постулату о том, что каждый ген имеет свой постоянный хромосомный локус. Сама МакКлинток даже получает обидное прозвище crazy Barbara (сумасшедшая Барбара). Однако позднее выясняется, что подобные мобильные генетические элементы присутствуют у всех живых организмов (стоит также упомянуть, что МакКлинток спустя 30 лет после своего открытия удостаивается Нобелевской премии в области физиологии и медицины [3]).
У животных, а конкретно, у дрозофилы, мобильные элементы впервые обнаруживают в лабораториях Хогнесса в США и Георгиева в СССР. Причем очень быстро становится ясно, что таких элементов огромное множество, в геномах они представлены очень широко, а по своим структурным и функциональным особенностям могут отличаться очень сильно. Изучение структуры различных классов мобильных элементов генома (МГЭ) приводит ученых к выводу об их родстве с вирусами. Жизненные циклы вирусов и многих МГЭ очень похожи, да и белки, кодируемые их генами, выполняют одни и те же функции, что отчетливо указывает на общность происхождения этих примитивных живых систем.
Экология генома: молекулярные паразиты и эндосимбионты
До 80-х — 90-х годов прошлого столетия тезис о том, что внутри человеческого генома могут находиться последовательности, собственно к человеку не имеющие никакого отношения, звучал бы дико. Однако сейчас мы сталкиваемся с тем, что геномы едва ли не всех эукариот кишат молекулярными паразитами! Известно, что подавляющее большинство вирусов могут встраивать свои ДНК в геном организма, который они заражают. При этом задача вируса очевидна — это размножение. Однако этой цели вирусы могут добиваться разными способами. С одной стороны, логично сразу же после встраивания начать синтезировать свои собственные белки, реплицировать свой генетический материал, а потом выходить из одной клетки организма и начинать заражать другие. В этой ситуации возникает небольшая проблема: клетка-хозяин погибает, а вслед за одной клеткой может погибнуть и весь организм, не дав возможности вирусу заразить другие организмы. Вступает в силу один из законов экологии: паразит, губящий своего хозяина, губит и себя самого. Поэтому возникает принципиально иная стратегия поведения молекулярных паразитов. После интеграции в геном многие вирусы начинают вести себя так, будто являются его неотъемлемой частью. Они не начинают активно копировать свой генетический материал, а остаются в сайте интеграции в виде так называемого провируса. Такой подход позволяет вирусам копировать информацию своего генома вместе с репликацией генома организма-хозяина, передаваясь по наследству потомкам этого хозяина. Британский генетик Джон Брукфилд (John F. Brookfield) в своих обзорах последних лет обосновывает введенный недавно термин «экология генома» [4], рассматривая в том числе и геном человека, как среду обитания всевозможных эндогенных ретровирусов, транспозонов и коротких сателлитных последовательностей. Причем между организмом-хозяином и такими сожителями ведется борьба по всем законам экосистем: паразит стремится размножиться, причиняя минимум беспокойства хозяину, а хозяин стремится либо паразита нейтрализовать, либо заставить работать на себя. При этом случаи мутуалистических отношений (сожительства с обоюдной пользой) между чужеродными последовательностями и геномом не редки. Одним из красивых примеров такого симбиоза является участие белковых продуктов эндогенных ретровирусов человека в процессе образования ткани плаценты. К тому же, широко известно, что относительно безопасные «одомашненные» вирусы могут препятствовать проникновению в клетки агрессивных вирусов извне.
Разумеется, помимо пользы от мобильных генетических элементов можно вполне ожидать и проблем. В частности, они могут провоцировать хромосомные аберрации, вызывать своими перемещениями мутации и изменения в активности генов, приводить к дестабилизации структуры всего генома. Взаимодействие между МГЭ и хозяйским геномом могут приводить к самым разнообразным и любопытным последствиям: от возникновения наследственных заболеваний до провоцирования процессов видообразования и образования новых генов.
Запутанная молекулярная инструкция
Проект «Геном человека» стартовал в начале девяностых и к настоящему времени завершен. Первые данные о составе нашего генома были опубликованы еще в 2001 году [5]. Тогда стало ясно, что на долю структурных генов (генов, содержащих информацию о строении белков или РНК) приходится около 5% всего генома (а ведь еще совсем недавно считалось, что кроме них в генетическом материале клетки быть ничего не должно). Самих же генов всего порядка 25–30 тысяч, что совсем не так много, как считалось. На долю же всевозможных мобильных последовательностей отводится целых 45% геномной ДНК! Остальное представлено повторами, поломанными неактивными генами и прочими техническими последовательностями. Причем все это невероятное разнообразие молекулярных текстов из поколения в поколение взаимодействует друг с другом, перестраивается, меняется местами и чуть ли не противоречит друг другу. Выяснить, как вся эта биоинформатическая каша из отдельных слов, написанных четырьмя молекулярными буквами-нуклеотидами, определяет внешний вид, характер и прочие особенности человека, — это задача, на решение которой уйдет еще очень много времени.
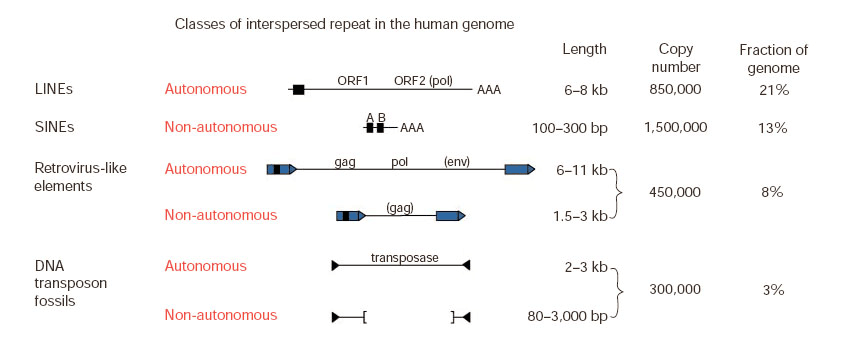
Рисунок 2. Классы повторов в геноме человека.
LINE — Long interspersed nuclear repeats. Одни из самых древних элементов. Содержат ген обратной транскриптазы и способны вносить разрывы в геномную ДНК при транспозиции. Часто образуют несовершенные копии.
SINE — Short interspersed nuclear repeats. Короткие последовательности, содержащие промотор полимеразы III. Их транспозиции происходят за счет белков, кодируемых генами LINE-элементов.
LTR (long terminal repeat) retrotransposons — группа элементов, по своей организации больше всего напоминающая вирусы (если точнее — ретровирусы). Считается, что часть ретротранспозонов произошла от вирусов, когда-то проникнувших в геном. Некоторые LTR-элементы сохраняют возможность покидать клетку-хозяина и инфицировать другие клетки. Включают от одного до нескольких генов.
DNA transposons — мобильные элементы, не требующие стадии образования РНК-копии для транспозиций. Кодируют фермент транспозазу, необходимую для перемещения.
Разумеется, значение проекта «Геном человека» сложно переоценить. Уже сейчас сделано множество интереснейших работ, которые были бы невозможны без прочтения последовательности человеческой ДНК. Однако каких-то 5–10 лет назад казалось, что секвенирование генома решит сразу все проблемы биологической науки, а заодно и медицины. В прессе было очень модно обсуждать прочтение «книги жизни», в которой содержится подробная инструкция о том, как собрать человеческий организм. Вроде бы оставалось лишь последовательно вникнуть в смысл каждой главы. И вдруг столь значительная книга, страницы которой так мечтали пролистать ученые конца XX века, на поверку оказывается зачитанным глянцевым журналом, пестрящим рекламой, вырезанными страницами, частными объявлениями с грамматическими ошибками и чьими-то заметками на полях. Найти что-то полезное в таком молекулярном издании довольно сложно, однако ученым не просто предстоит отделить в нем ценную информацию от бессмыслицы, но также понять алгоритм того, как полезные отрывки сочетаются и дополняют друг друга, реализуясь в целостной картине человеческого организма. И это не самая простая головоломка.
Литература
- James D Watson, Francis Harry Compton Crick. (2007). THE CLASSIC: Molecular Structure of Nucleic Acids. Clinical Orthopaedics and Related Research. 462, 3-5;
- McClintock B. (1950). The origin and behavior of mutable loci in maize. Proc. Natl. Acad. Sci. U.S.A. 36, 344–355;
- Нобелевские лауреаты. Барбара МакКлинток. Электронная библиотека «Наука и техника»;
- John F. Y. Brookfield. (2005). The ecology of the genome — mobile DNA elements and their hosts. Nat Rev Genet. 6, 128-136;
- Eric S. Lander, Lauren M. Linton, Bruce Birren, Chad Nusbaum, Michael C. Zody, et. al.. (2001). Initial sequencing and analysis of the human genome. Nature. 409, 860-921;
- Orgel L.E. and Crick F.H. (1980). Selfish DNA: the ultimate parasite. Nature. 284, 604–607;
- F. P Ryan. (2004). Human endogenous retroviruses in health and disease: a symbiotic perspective. JRSM. 97, 560-565.



Комментарии
1Чтобы оставить комментарий, необходимо
войти