От искры к пожару: как рождается и путешествует нервный импульс
05 марта 2026
От искры к пожару: как рождается и путешествует нервный импульс
- 688
- 0
- 4
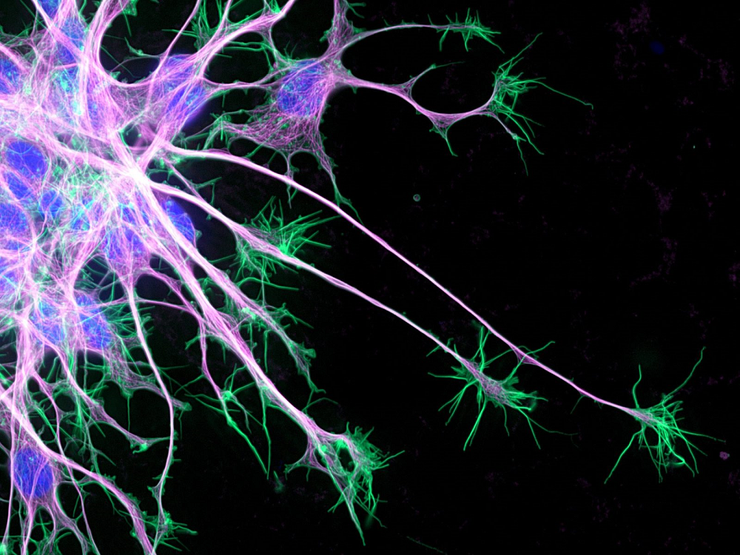
Изображение растущих нервных клеток с конкурса Nikon’s Small World 2019. При помощи специальных флуоресцентных красителей можно увидеть микротрубочки, растущие внутри увеличивающихся нейрональных отростков (розовые). Актиновые филаменты (зеленые) при этом тянутся дальше в обратную от тел нейронов сторону. Ядра окрашены в синий цвет.
Рисунок в полном размере.
-
Автор
-
Редакторы
Статья на конкурс «Био/Мол/Текст»: Что общего у мгновенного отдергивания руки от горячего и внезапно вспыхнувшей в голове гениальной мысли? Их рождает один и тот же удивительный механизм — нервный импульс. Это не просто возбуждение, которое клетки передают друг другу, а настоящий ток — уникальный электрохимический механизм, который возникает, словно вспышка, и бежит по нервному волокну.
В этой статье мы проследим весь путь этого импульса: как нейрон создает и поддерживает потенциал покоя, что такое порог возбуждения — тот самый критический рубеж, за которым рождается импульс, и как мембранные каналы обеспечивают идеально скоординированный сценарий деполяризации и реполяризации. Вы узнаете, как именно импульс передвигается по волокнам и зачем для этого нужен миелин. А самое главное — как один маленький нейрон передает эстафету другим клеткам, чтобы мы могли слаженно управлять своим телом.

Конкурс «Био/Мол/Текст»-2025/2026
Эта работа опубликована в номинации «Свободная тема» конкурса «Био/Мол/Текст»-2025/2026.
Генеральный партнер конкурса — международная инновационная биотехнологическая компания BIOCAD.
Партнер номинации — компания SkyGen: передовой дистрибьютор продукции для life science на российском рынке.
«Книжный» спонсор конкурса — «Альпина нон-фикшн»
Два химических мира: зачем нейрону изолированная мембрана?
Нервные клетки — сборное понятие. Они сильно различаются по размеру, форме, количеству отростков и выполняемым функциям. Однако глобально все клетки нервной ткани подразделяются на две основные ветви. Принято выделять нейроны (клетки, способные возбуждаться и проводить импульс) и нейроглию (клетки, которые помогают и поддерживают нейроны в этом процессе). Более подробно про виды нервных клеток можно почитать в следующих статьях на «Биомолекуле»: «Ликбез по ЦНС» [1] и «Микроглия: роль „иммунных“ клеток центральной нервной системы в здоровом мозге и при нейродегенеративных заболеваниях» [2].
Нейрон — это высокоспециализированная клетка с уникальной архитектурой, где каждый элемент идеально приспособлен для конкретной функции (рис. 1). Рассмотрим ключевые компоненты:

Рисунок 1. Строение нейрона. На схеме обозначены основные части нервной клетки: тело (сома) с дендритами, аксон с миелиновой оболочкой, образованной клетками Шванна, и специализированные участки — перехваты Ранвье, обеспечивающие сальтаторное проведение импульса. Терминали аксона предназначены для передачи сигнала другим клеткам.
- тело клетки (сома) — своего рода «центр управления». Здесь расположены ядро и основные органеллы, обеспечивающие жизнедеятельность нейрона;
- дендриты — многочисленные древовидные «антенны», отходящие от тела клетки. Их основная задача — принимать входящие сигналы от тысяч других нейронов через специальные контакты, известные как синапсы;
- аксон — единственный длинный и тонкий «кабель передачи», который может тянуться на расстояние до метра и более. Например, аксоны, идущие от спинного мозга к кончикам пальцев, обеспечивают проведение электрических импульсов без затухания на значительные расстояния;
- клеточная мембрана — это тонкая пленка, которая состоит из липидного бислоя, пронизанного специализированными белками. Эти белки образуют каналы, с помощью которых клетка обменивается веществами с окружающей средой. Она разделяет внутреннее пространство нейрона от внешней среды и служит избирательным барьером (рис. 2).
Именно мембрана выступает главной причиной и организатором возникновения нервного импульса.

Рисунок 2. Строение мембраны. Мембрана представлена в виде липидного бислоя, где полярные гидрофильные «головки» обращены к водной среде (цитозолю и межклеточной жидкости), а гидрофобные «хвосты» образуют внутренний слой. Белки, встроенные в мембрану (например, ионный канал), обеспечивают ее специализированные функции.
Важно понимать: пространства внутри и снаружи клетки — это два разных химических мира, каждый со своим уникальным составом. Поэтому для клетки жизненно важно контролировать, какие вещества проникают внутрь, а какие покидают ее. Эта способность обеспечивается уникальным свойством клеточной мембраны — избирательной проницаемостью, которая достигается за счет работы мембранных белков.
Без участия белковых каналов, напрямую через липидный слой, может проникнуть лишь очень ограниченный набор веществ — небольшие незаряженные липофильные молекулы. Для транспорта всех остальных необходима работа специальных транспортных белков (рис. 3).

Рисунок 3. Виды мембранного транспорта. Иллюстрирует пути перемещения веществ: простая диффузия, транспорт через ионные каналы (управляемые и неуправляемые) и активный транспорт. Эти процессы лежат в основе формирования и поддержания мембранного потенциала и проведения импульса.
рисунок автора статьи
Транспорт веществ через клеточную мембрану может идти двумя принципиально разными путями: по градиенту концентрации (из области, где вещества больше, туда, где его меньше) или против него. От этого фундаментального различия зависит тип используемого клеткой мембранного транспорта. Для перемещения веществ по градиенту существуют специальные белки, формирующие каналы. Они бывают двух основных типов:
- «неуправляемые» каналы — это постоянно открытые «проходы», своеобразные поры, через которые пассивный обмен ионами или молекулами идет непрерывно. К таким каналам относятся, например, калиевые каналы утечки, которые есть во всех клетках;
- «управляемые» каналы — это настоящие молекулярные «ворота», которые клетка открывает только в ответ на определенный сигнал (например, изменение электрического поля или связывание с конкретной молекулой). Так, изменение электрического поля открывает натриевые и калиевые потенциал-зависимые каналы.
Такой вид транспорта использует готовую движущую силу — разницу концентраций — и не требует от клетки прямых энергозатрат, что позволяет ей экономить ресурсы. Однако, когда вещество необходимо перенести против градиента концентрации (например, накачать ион внутрь клетки, хотя его и так там много), клетке приходится тратить энергию. Этот процесс, известный как активный транспорт, осуществляется специальными белками-насосами, которые для этого расщепляют молекулы АТФ.
От покоя к вспышке: как в нейроне рождается импульс
Разная концентрация ионов на разных сторонах клеточной мембраны порождает не только химический, но и электрический дисбаланс. В итоге внутренняя поверхность мембраны приобретает отрицательный заряд по сравнению с внешней средой, где преобладает положительный заряд.
Эта разница в зарядах создает мембранный потенциал — особенность, характерная для всех клеток организма. Но именно в возбудимых клетках, особенно в нейронах, критически важен потенциал покоя — стабильный отрицательный заряд около −70 мВ, сохраняющийся, когда нейрон находится в состоянии бездействия [3].
Основную роль в поддержании этого потенциала играет натрий-калиевая АТФ-аза — молекулярный насос, встроенный в мембрану и функционирующий против градиентов концентрации. Непрерывно выводя из клетки три иона натрия (Na+) и впуская два иона калия (K+), этот насос совершает работу, требующую затрат энергии за счет расщепления АТФ. Именно благодаря такому асимметричному обмену не только сохраняется разница в концентрации ионов, но и формируется отрицательный заряд внутри клетки.
Мы рассмотрели, как клетка создает и поддерживает свой мембранный потенциал. Так как же это связано с возникновением импульса в нейронах?

Рисунок 4. Фазы потенциала действия. График иллюстрирует изменение мембранного потенциала во времени в ответ на стимул. Показан переход от потенциала покоя (-70 мВ) к деполяризации: подпороговые локальные ответы суммируются, и при достижении критического уровня (-55 мВ) активируются потенциал-управляемые натриевые каналы. Это вызывает лавинообразный вход Na+ — фазу быстрой деполяризации (фаза нарастания). Пик потенциала действия (около 0 мВ) соответствует инактивации Na+-каналов и активации K+-каналов, что приводит к реполяризации и следовым потенциалам. Демонстрируется фундаментальный принцип возбудимых мембран — закон «всё или ничего».
Мы оставили наш нейрон в состоянии идеально организованного напряжения — живой микроскопической батарейкой с зарядом в −70 мВ. Но какой смысл в том, чтобы быть батарейкой, если никогда не разряжаться? Энергию нужно использовать. Стабильный потенциал покоя — это не конечная цель, а идеальная стартовая площадка для генерации нервного импульса.
Тишина потенциала покоя обманчива. Она длится ровно до того момента, пока на дендриты не поступит достаточно сильный сигнал. Этот сигнал вызывает небольшое локальное возмущение, которое должно преодолеть критический рубеж, чтобы из простого возмущения родилась та самая «вспышка» — потенциал действия (рис. 4).
- Исходное состояние. В спокойном состоянии мембрана нейрона поляризована, а ее потенциал покоя стабильно держится возле отметки −70 мВ. Работа натрий-калиевого насоса создала четкие ионные градиенты: снаружи доминируют ионы Na+, внутри — K+. Потенциал-зависимые натриевые каналы в основном закрыты, но находятся в состоянии боевой готовности. Одновременно через калиевые каналы утечки потихоньку выходят ионы K+, создавая небольшую проницаемость мембраны. В результате нейрон напоминает заряженную батарейку, терпеливо ожидающую команды.
- Запускающий стимул и локальный ответ. Когда к нейрону приходит возбуждающий сигнал (скажем, через синапс ), возникает легкая деполяризация — мембранный потенциал немного снижается (например, с −70 мВ до −65 мВ). Это происходит из-за открытия лиганд-зависимых рецепторных каналов и притока положительно заряженных ионов. Такой сдвиг носит локальный и плавный характер. Если стимул слишком слаб, возбуждение просто затухает без последствий. Но когда деполяризация достигает критического порога (примерно −55 мВ), начинается необратимая цепная реакция, приводящая к рождению нервного импульса.
- Фаза деполяризации. Порог — это та самая точка невозврата. При его достижении срабатывает ключевой механизм:
- Массовое открытие натриевых каналов. Деполяризация до −55 мВ — это долгожданный сигнал для потенциал-зависимых Na+-каналов. Их молекулярные «ворота» распахиваются, пропуская внутрь клетки настоящий ионный поток. Ионы натрия, которых снаружи накопилось очень много, теперь лавинообразно устремляются в клетку. Движут их сразу две силы: химический градиент (стремление из области высокой концентрации в низкую) и электрический градиент (их манит отрицательный заряд внутри нейрона).
- Мембранный потенциал стремительно стремится к нулю и далее — к равновесному потенциалу для натрия, достигая пика около +30 мВ. Это фаза стремительного «взлета» на графике ПД.
- Фаза реполяризации. Достигнув пика примерно на отметке +30 мВ, потенциал действия запускает следующий этап — процесс восстановления исходного электрического состояния клетки. Этот механизм включает три последовательных события:
- Блокировка натриевых каналов. Резкий скачок потенциала заставляет потенциал-зависимые Na+-каналы захлопнуться. Срабатывают специальные инактивирующие «ворота» в структуре белка, надежно перекрывая ионный путь. Любопытно, что каналы теперь переходят не просто в закрытое состояние, а в особое инактивированное — своего рода временный молекулярный арест, делающий их неспособными к открытию.
- Активация калиевых каналов. Пока натриевые каналы блокируются, деполяризация мембраны будит к жизни более неторопливых собратьев — потенциал-зависимые K+-каналы. Их запаздывающее открытие создает настоящий прорыв: ионы калия мощным потоком устремляются из клетки наружу, следуя по электрохимическому градиенту.
- Восстановление мембранного потенциала. Массовый выход положительно заряженных ионов калия быстро смещает заряд внутренней среды клетки в отрицательную область. Этот процесс, наблюдаемый на графике как крутой спад кривой после пика, обеспечивает возврат мембранного потенциала к значениям, близким к потенциалу покоя.
- Рефрактерный период. Любопытно, но сразу после рождения импульса нейрон ненадолго становится «глухим» к новым сигналам. Это состояние — рефрактерный период — своеобразный физиологический тайм-аут, и протекает он в две стадии.
- Абсолютный рефрактерный период — время полной невосприимчивости. Он захватывает пик деполяризации и почти всю реполяризацию. В чем его причина? Натриевые каналы в этот момент не просто закрыты, а структурно инактивированы. Представьте дверь, не только запертую на ключ, но и с заклинившим механизмом — открыть ее невозможно, какой бы мощный стимул ни действовал. Именно это и гарантирует, что импульс бежит только вперед, не имея шансов повернуть вспять.
- Относительный рефрактерный период наступает следом. Мембранный потенциал в эту фазу часто «перескакивает» отметку покоя, становясь еще более отрицательным (явление следовой гиперполяризации, около −75 мВ). Виной тому все еще открытые калиевые каналы, через которые продолжается утечка положительных зарядов. В это время нейрон уже можно «расшевелить», но для этого потребуется стимул заметно сильнее обычного.
Миелин и сальтаторное проведение — скоростная магистраль для нервного импульса
Наш потенциал действия (ПД) продолжает свое путешествие по мембране аксона. Но представьте на мгновение: если бы импульсу приходилось последовательно возбуждать каждый микрон гигантского — длиной до метра! — аксона, процесс оказался бы чудовищно медленным и неэффективным!
К счастью, нейроны нашли элегантное решение этой проблемы, создав настоящую систему биологических «скоростных магистралей».
Ключевую роль здесь играет миелин — многослойная липопротеиновая оболочка, напоминающая изоляцию электрического кабеля. Она не просто плотно оборачивает аксон, но и формирует прерывистое покрытие. Между миелиновыми сегментами остаются микроскопические оголенные участки — перехваты Ранвье, которые становятся критически важными точками для генерации сигнала.
И здесь в игру вступают два типа клеток нейроглии (рис. 5):

Рисунок 5. Олигодендроциты и шванновские клетки. Иллюстрация демонстрирует ключевое различие в формировании миелина: в периферической нервной системе (ПНС) одна клетка Шванна миелинизирует сегмент одного аксона, тогда как в центральной нервной системе (ЦНС) один олигодендроцит может образовывать миелиновые оболочки вокруг нескольких аксонов одновременно. Миелин служит изолятором, значительно увеличивающим скорость проведения нервного импульса.
- Шванновские клетки — это часть периферической нервной системы (нервы, идущие к мышцам и органам). Каждая такая клетка формирует один миелиновый сегмент вокруг одного аксона.
- Олигодендроциты — их «коллеги» в центральной нервной системе (головном и спинном мозге). Причем один олигодендроцит может «взять шефство» сразу над несколькими аксонами, формируя миелиновые сегменты на каждом из них.
Так в чем же заключается секрет высокой скорости? Все дело в удивительном механизме сальтаторного проведения — то есть скачкообразного. Мы уже знаем, что потенциал действия рождается благодаря работе потенциал-зависимых ионных каналов. Оказывается, эти каналы сосредоточены в основном в перехватах Ранвье. А вот под надежной миелиновой изоляцией их практически нет.
Представьте себе этот процесс:
- Потенциал действия возникает в первом перехвате Ранвье.
- Возникающий ионный ток не растекается вдоль мембраны — миелиновая оболочка, обладающая высоким сопротивлением, мешает этому. Вместо этого заряд практически мгновенно «перепрыгивает» к следующему перехвату, подобно электрическому току по проводу.
- Этого скачка оказывается достаточно для деполяризации мембраны в новом перехвате до критического уровня. Срабатывает цепная реакция — лавинообразно открываются натриевые каналы, и рождается следующий полноценный потенциал действия.
- Процесс повторяется снова и снова. Импульс, словно искра по бикфордову шнуру, перескакивает от одного перехвата к другому, не тратя времени на участки под миелином.
Такой подход дает нейрону два ключевых преимущества:
- скорость — проведение сигнала по миелинизированным волокнам достигает 120 м/с — в десятки раз быстрее, чем по волокнам без миелина;
- энергоэффективность — нейрону не нужно генерировать потенциал действия на каждом микроне аксона, а только в перехватах, что существенно экономит энергию (ионы Na+ и K+).
Наш импульс, начавшись скромным электрическим всплеском в теле нейрона, преображается в стремительную волну, летящую вдоль аксона от одного перехвата к другому. Но что же происходит, когда этот сигнал достигает конечного пункта своего маршрута — кончика аксона? Каким образом он передает информацию следующему нейрону, мышечному волокну или железе? Перед нами разворачивается заключительная глава этой удивительной истории — загадочный процесс синаптической передачи.
Химический мост: что такое нейромедиаторы и как они передают сообщения
Преодолев весь аксон на огромной скорости, нервный импульс наконец достигает своей конечной цели — специального окончания, где происходит передача информации следующей клетке. Этот контакт, известный как синапс (подробнее об этом читайте на «Биомолекуле»: [1], [4], [5]), становится местом, где электрический язык потенциалов действия преобразуется в универсальный химический код, понятный другим клеткам.
Между двумя нервными клетками существует крошечный промежуток — синаптическая щель. Нейрон, передающий сигнал, называют пресинаптическим, а принимающий — постсинаптическим [4].
Но как именно это происходит? Когда потенциал действия добирается до пресинаптического окончания, разворачивается настоящий молекулярный фейерверк.
Первый шаг — кальциевый сигнал. Достигнув конечного участка аксона, электрический импульс деполяризует его мембрану. Это мгновенно открывает потенциал-зависимые кальциевые каналы. Ионы кальция (Ca2+), которых снаружи значительно больше, устремляются внутрь, запуская следующий этап.
Далее начинается химическая стадия. В пресинаптическом окончании находятся многочисленные синаптические пузырьки с нейромедиаторами — особыми химическими веществами-посредниками. Кальциевый всплеск заставляет эти пузырьки перемещаться к мембране, сливаться с ней и высвобождать свои запасы в синаптическую щель.
Путешествие через разрыв. Молекулы медиатора быстро диффундируют через узкую щель (это занимает доли миллисекунды) и достигают мембраны принимающего нейрона.
Молекулярное узнавание. На поверхности постсинаптического нейрона расположены специальные рецепторные белки, которые работают как замки для медиаторных ключей. Когда молекула медиатора связывается с рецептором, происходит изменение его пространственной структуры, что приводит к открытию связанных с ним ионных каналов.
Рождение нового сигнала. В зависимости от типа открывшихся каналов в принимающем нейроне возникает один из двух вариантов ответа:
- Возбуждающий постсинаптический потенциал (ВПСП) — когда открываются каналы для положительных ионов (например, Na+). Их вход вызывает небольшую деполяризацию, сдвигая мембранный потенциал ближе к порогу возбуждения.
- Тормозной постсинаптический потенциал (ТПСП) — если открываются каналы для отрицательных ионов (Cl⁻) или выхода положительных ионов (K+). Это вызывает гиперполяризацию, временно снижая возбудимость клетки.
Финальный этап — интеграция. Каждый нейрон постоянно обрабатывает тысячи таких возбуждающих и тормозных сигналов. Ключевая задача клетки — просуммировать все эти ВПСП и ТПСП. Если общий эффект достигает критического порога в области аксонного холмика, рождается новый потенциал действия, и весь цикл повторяется. Если же порог не достигнут, импульс не возникает, и информация не передается дальше.
Заключение
На этом завершается удивительный путь нервного импульса. От хрупкого равновесия потенциала покоя — через мощную вспышку потенциала действия — к стремительному движению по миелиновым «магистралям» и, наконец, к тонкому химическому диалогу в синаптической щели.
Поражает, что этот сложнейший механизм, повторяющийся миллиарды раз каждую секунду, составляет основу всей нашей жизнедеятельности. Обычное движение рукой и глубокая мысль, мимолетная эмоция и творческая идея — все это рождается благодаря слаженной работе ионов, мембранных каналов и молекул-медиаторов.
Именно этот процесс, напоминающий то ли идеально отлаженный механизм, то ли виртуозный танец, определяет саму возможность нашего существования и восприятия мира.
Литература
- Ликбез по ЦНС;
- Микроглия: роль «иммунных» клеток центральной нервной системы в здоровом мозге и при нейродегенеративных заболеваниях;
- Формирование мембранного потенциала покоя;
- Как происходит выделение нейромедиатора;
- Мозг, общение нейронов и энергетическая эффективность;
- Холл Дж. Э. Медицинская физиология. 3-е изд., испр. и доп. — М.: «Логосфера», 2024. — 1344 с.;
- Коган А. Б. Основы физиологии высшей нервной деятельности. 2-е изд., перераб. и доп. — М.: «Высшая школа», 1988. — 368 с..



Комментарии
0Чтобы оставить комментарий, необходимо
войти