Собственной персоной: ученым удалось вырастить в лаборатории вероятного предка эукариот
12 сентября 2019
Собственной персоной: ученым удалось вырастить в лаборатории вероятного предка эукариот
- 1443
- 0
- 7
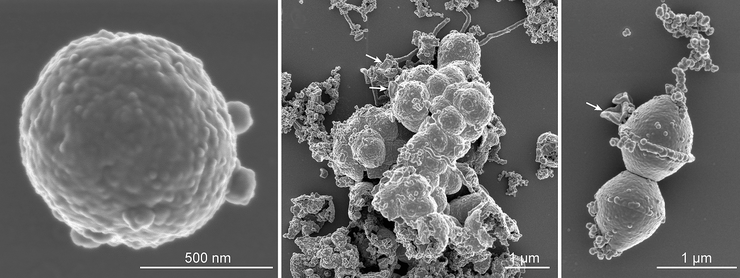
Микрофотографии Candidatus Prometheoarchaeum syntrophicum strain MK-D1, полученные с помощью сканирующей электронной микроскопии. Слева — одиночная клетка; в центре — скопление клеток, покрытых полисахаридным чехлом; справа — делящаяся клетка с цепочкой внеклеточных везикул. Фотография в полном разрешении.
-
Автор
-
Редактор
Происхождение эукариот было и остается одной из интереснейших проблем эволюционной биологии. В 2015 году в Nature появилось сообщение, что с помощью метагеномики удалось получить последовательность генома археи ранее неизвестной группы, причем в этом геноме удалось найти гены, которые ранее считались уникальными для эукариот (в частности, ген, близкий к кодирующему актин). Более того, филогенетический анализ показал, что археи новооткрытой группы, получившей название Lokiarchaeota, и эукариоты образуют монофилетическую кладу (группу организмов с общим предком), и эукариоты, вероятно, произошли от локиархеот. Недавно японские ученые сообщили, что им удалось в лабораторных условиях вырастить культуру археи из типа Lokiarchaeota, которую они назвали Candidatus Prometheoarchaeum syntrophicum strain MK-D1. Какие загадки происхождения эукариот мы сможем разрешить, имея на руках чистую культуру их непосредственных предков, а не абстрактный геном? Попробуем разобраться.
Подавляющее большинство микроорганизмов, обитающих на нашей планете, в принципе невозможно вырастить в лаборатории ввиду их очень нетипичных физиологических потребностей, которым не удовлетворяют существующие лабораторные среды. Кроме того, зачастую микробы образуют тесные метаболические связи друг с другом и не могут быть выращены по отдельности. Наконец, многие микробы размножаются так медленно, что даже при использовании оптимальной среды на получение культуры, пригодной для изучения, может уйти несколько лет. Изучать такие организмы зачастую можно лишь в их природных местообитаниях. Впрочем, благодаря развитию метагеномики стало возможным получать последовательности геномов организмов из образцов окружающей среды, собранных в самых разных местах земного шара, однако зачастую голая нуклеотидная последовательность не может дать исчерпывающую информацию о биологии микроорганизма.
Именно поэтому сообщение о выделении в лабораторных условиях живых архей, являющихся вероятными предками эукариот, стало настоящей сенсацией. Японские ученые разместили в репозитории препринтов bioRxiv публикацию, в которой утверждают, что в результате десятилетней работы им удалось получить культуру археи группы Lokiarchaeota — вероятного предка эукариот [1], [2]. Исследователи дали микроорганизму сложное название Candidatus Prometheoarchaeum syntrophicum strain MK-D1. В настоящее время Lokiarchaeota вместе с двумя другими недавно выделенными типами архей, Heimdallarchaeota и Odinarchaeota, объединяются в группу Asgard. Археи этой группы очень близки к эукариотам и, согласно полученным с помощью метагеномики геномным последовательностям их представителей, обладают многими чертами, ранее считавшимися уникальными для эукариот.
Получение культуры столь интересных для эволюционных биологов архей стало сложнейшим испытанием, продолжавшимся более десяти лет. Изначально ученые взяли в работу образец морских донных отложений с большой глубины. Для дальнейшей культивации попавших в образец микробов разработали специальный биореактор с непрерывной подачей метана, в котором поддерживались анаэробные условия. Спустя долгие 2000 дней культивации плотность микроорганизмов в образце достигла пригодных для изучения значений. В получившейся «сборной солянке» оказалось множество неродственных друг другу микроорганизмов, которые до этого не культивировались в лаборатории. В их числе были и археи группы Asgard.
Для обогащения смешанной культуры и ее разделения на чистые культуры образцы из биореактора перенесли в стеклянные трубки, содержащие среду с простыми субстратами. Спустя год культивации в одной из трубок, где среда имела температуру 20 °C и содержала, помимо казаминовых кислот, четыре антибиотика, подавляющие рост бактерий, исследователи обнаружили слабое замутнение. Анализ генов рибосомной РНК, входящей в состав малой субъединицы рибосом, показал, что в трубке обитает простое сообщество, состоящее из архей родов Halodesulfovibrio и Methanogenium, а также небольшого количества представителей Lokiarchaeota. Чтобы отделить локиархей от их спутников, ученые готовили несколько последовательных субкультур: когда плотность локиархей в предшествующей культуре достигала максимальных значений, разводили небольшой ее объем в новой среде и продолжали инкубацию. После шести пересевов доля локиархеот, которые и получили название Candidatus Prometheoarchaeum syntrophicum strain MK-D1 (здесь и далее для краткости мы будем называть их MK-D1), в культуре достигла 13%, при этом другие 85% клеток относились к археям рода Halodesulfovibrio, а оставшиеся 2% составляли археи рода Methanogenium. Как показало исследование культуры с помощью флуоресцентной гибридизации in situ и сканирующей электронной микроскопии, археи всех трех родов находились в тесном физическом контакте друг с другом и обменивались разнообразными веществами, в частности, водородом. С помощью дальнейших пересевов исследователи смогли получить культуру, полностью лишенную Halodesulfovibrio и представляющую собой почти чистую культуру MK-D1 (с небольшой примесью архей из рода Methanogenium). Суммарно путь от образца морского грунта до получения чистой культуры занял целых 12 лет!
MK-D1 оказались чрезвычайно медленно растущими археями с временем удвоения от 14 до 25 дней. Эти мелкие кокки диаметром в среднем 550 нм формируют скопления, окруженные полисахаридным чехлом. Детальное изучение клеток с помощью различных видов микроскопии показало, что каких-либо структур, похожих на органеллы, в них не содержится, хотя от клеток MK-D1 отпочковываются мембранные везикулы. Самым необычным в морфологии MK-D1 оказалось то, что они формируют мембранные выросты (протрузии) различной длины и диаметром от 80 до 100 нм (рис. 1). В отличие от протрузий, описанных у других архей [3], протрузии MK-D1, как оказалось, могут обильно ветвиться, причем количество протрузий значительно увеличивалось, когда клетки проходили позднюю экспоненциальную фазу роста.

Рисунок 1. Протрузии клеток MK-D1: ветвящиеся (слева) и неветвящиеся (справа)
С точки зрения метаболизма MK-D1 оказались способны к анаэробному окислению десяти аминокислот, причем в процессе окисления они продуцировали водород и муравьиную кислоту. Эти вещества, накапливаясь, подавляют рост MK-D1, поэтому в природе MK-D1 обитают только в непосредственном контакте с потребляющими муравьиную кислоту и водород партнерами — археями из рода Methanogenium.
Наличие у MK-D1 необычных клеточных выростов, способных к ветвлению, позволило ученым предложить новую модель возникновения эукариот (эукариогенеза), которую они назвали Entangle—Engulf—Enslave (E3) («обхватить—проглотить—поработить»). Накопление в атмосфере кислорода, связанное с бурной деятельностью цианобактерий, поставил остальных микробов, не имеющих защиты от кислорода, перед выбором: или полностью уйти в анаэробные среды, или научиться самим нейтрализовать кислород, или же завести внутриклеточного эндосимбионта, способного обезвреживать кислород (который потом и дал начало митохондриям эукариот). Разные археи группы Asgard пошли разными путями. Heimdallarchaeota приобрели способность к самостоятельному анаэробному дыханию, то есть стали сами обезвреживать кислород, а предки эукариот — локиархеи — «поработили» бактерий, способных его нейтрализовать (рис. 2). Вероятно, мощные клеточные выросты, выявленные у MK-D1, способствовали такому захвату эндосимбионтов.
О сложных взаимоотношениях доисторических бактерий и архей читайте в статье «Закинули археи эволюционный невод и вытянули...» [4]. — Ред.

Рисунок 2. Филогенетическое дерево, иллюстрирующее положение MK-D1 среди архей и эукариот
Литература
- Imachi H., Nobu M.K., Nakahara N., Morono Y., Ogawara M., Takaki Y. et al. (2019). Isolation of an archaeon at the prokaryote-eukaryote interface. bioRxiv;
- Anja Spang, Jimmy H. Saw, Steffen L. Jørgensen, Katarzyna Zaremba-Niedzwiedzka, Joran Martijn, et. al.. (2015). Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature. 521, 173-179;
- Археи «хамят» и помогают;
- Закинули археи эволюционный невод и вытянули....



Комментарии
0Чтобы оставить комментарий, необходимо
войти