Цели и вознаграждения искажают карту пространства в мозге
30 апреля 2019
Цели и вознаграждения искажают карту пространства в мозге
- 1803
- 0
- 8
Для ориентации в пространстве мы используем навигационную систему мозга, сформированную под воздействием целей и наград.
-
Автор
-
Редакторы
Специальные нейроны в мозге — клетки решетки в энторинальной коре, прилежащей к гиппокампу, — помогают животным ориентироваться в пространстве. Активность этих нейронов размечает окружающую среду гексагональной решеткой, выполняющей роль пространственной карты. Однако эта решетка вовсе не регулярная — она искривляется, если в пространстве есть цели и вознаграждения, как показали сразу две группы ученых из Стэнфорда и Австрийского института науки и технологий. Но даже на этом удивительные свойства гиппокампа и его соседей не заканчиваются: исследования показывают, что пространственные карты мозга могут участвовать в кодировании не только целей и вознаграждений, но и визуальных стимулов, а также более абстрактных когнитивных процессов, таких как социальные структуры, воспоминания и идеи.
Репутация «системы GPS-навигации» в мозге прочно закрепилась за гиппокампом и прилежащей к нему энторинальной корой (вместе их можно называть гиппокампальной формацией) после вручения Нобелевской премии по физиологии и медицине в 2014 году Джону О`Кифу и супругам Мозер. «Биомолекула» подробно писала об открытии врожденной системы ориентации в пространстве [1], включая нейроны места и решетки, которые, безусловно, стали фундаментом в понимании того, как мозг размечает пространство.
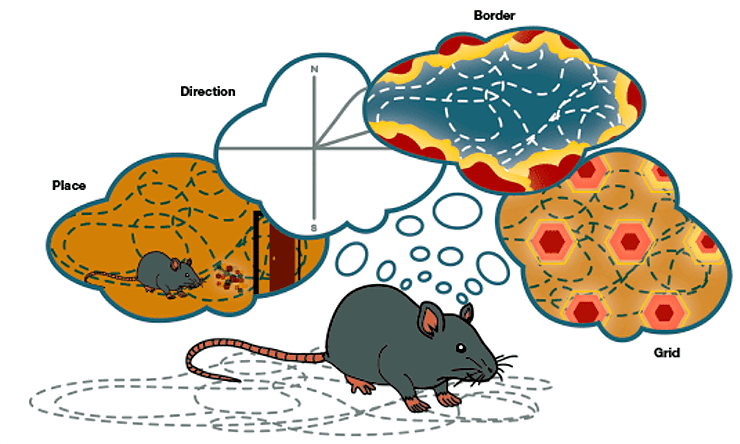
Нейроны места (place cells) в гиппокампе заметил О`Киф еще в семидесятых [2]: эти клетки были активны, когда животное (все ранние эксперименты проводили на крысах) находилось в определенном месте в пространстве — скажем, в левом дальнем углу лабиринта — и связывало это место с чем-то запоминающимся и локальным (рис. 1). У нейронов решетки (grid cells), описанных Мозерами [3] в области, тесно связанной с гиппокампом — энторинальной коре, — более сложный паттерн активности: они «загораются» во множественных местах пространства. При внимательном рассмотрении их поля активности складываются в гексагональную решетку, равномерно покрывающую пространство (рис. 1) [4]. Ученые называют эту решетку инвариантной — где бы не находилось животное, решетка сохраняет форму и помогает ориентироваться, хотя может изменять масштаб. Если испытуемый с завязанными глазами случайно бродит по комнате, то, возможно, он не сможет сказать, в каком месте остановился, так как не видит ориентиров, но ему довольно точно удастся предположить, какое расстояние он прошел и в каком направлении.

Рисунок 1. Разные типы нейронов, участвующие в навигации и найденные в гиппокампе и прилежащих районах мозга крысы. Слева направо: клетки места, активные в одной точке пространства; клетки решетки, имеющие несколько полей активности, организованные в форме гексагональной решетки; клетки границы, обозначающие геометрию пространства; клетки направления, указывающие на абсолютное направление головы.
Хотя нейроны решетки играют определяющую роль в интеграции пространственной информации, найдены и другие типы клеток, которые им в этом помогают. Клетки границы (border cells) активны тогда, когда животное находится у физической преграды (например, стены) и обозначают геометрию пространства. Клетки направления (head-direction cells) сигнализируют об абсолютном направлении головы животного и являются в истинном смысле компасом мозга (рис. 1). Группа Нахума Ульяновского в Институте Вайцмана, изучающая трехмерную пространственную ориентацию на летучих мышах, нашла в гиппокампе социальные клетки места (social place cells) [5], сообщающие, где в пространстве находится другая особь того же вида, — необходимое знание для социальных животных, задействованных в совместной навигации. Кроме этого, в гиппокампальной формации найдены нейроны, отмечающие скорость и определенную цель в пространстве.
Со стороны система координации разных нейронов, ответственных за ориентацию, выглядит изящной и сбалансированной. С одной поправкой. Все эксперименты проводили в маленьких и однородных пространствах (аренах или лабиринтах) размером в несколько метров. Никто не знает, как нейроны работают в природных условиях огромного размера с разнообразием опасностей, наград и поощрений.
На первый вопрос пытается ответить группа Нахума Ульяновского, которая не только подтвердила существование всех тех же типов клеток (места, решетки, направления и границы) у летучих мышей при ориентации в двухмерном и трехмерном пространстве, но и показала в предварительных экспериментах, что летучие мыши способны помнить маршруты длиной в десятки километров (рис. 2) [6]. Ученые отслеживали перемещения летучих мышей в естественной для них среде с помощью миниатюрных GPS-датчиков — одно из животных летало к одному и тому же фруктовому дереву в 15 километрах от пещеры на протяжении семи дней подряд (рис. 2а)! А если летучую мышь увозили за 44 км от ее пещеры и выпускали, она направлялась к пещере, когда была сыта, или прямиком к фруктовому дереву, когда была голодна (рис. 2б).

Рисунок 2. Эксперименты показывают, что летучие мыши запоминают расстояния величиной в десятки километров. а — Траектория движения одной летучей мыши, которая на протяжении семи дней летала к одному и тому же фруктовому дереву в 15 км от пещеры. б — Траектории полета двух летучих мышей, которых увезли за 44 км от пещеры и выпустили: в сытом состоянии они отправились к пещере (красный след), а в голодном — прямиком к столу (фруктовому дереву).
Если размер поля нейрона места в маленькой арене обычно 10–20 сантиметров, то на таком большом расстоянии наверняка происходит что-то интересное: либо нейроны места и решетки «масштабируют» свои поля, либо один нейрон места активен в нескольких маленьких полях, — никто не знает. Активность нейронов невозможно изучать на таком большом и неконтролируемом пространстве, поэтому специально для группы Ульяновского недалеко от Института Вайцмана строят специальный километровый экспериментальный тоннель для летучих мышей [7]. Благодаря ему удастся узнать, как система навигации работает «по-настоящему».
На второй вопрос — как мозг использует память и когнитивную информацию для преобразования пространственных карт — частично отвечают две одновременно вышедшие статьи в Science от разных групп: из Стэнфордского университета [8] и Австрийского института науки и технологий [9]. Их общая идея в том, что для эффективной и биологически релевантной навигации мозг должен не только надежно «размечать» пространство геометрически, но и встраивать в него непространственную информацию: цели и награды, такие как источники пищи. Обе статьи показывают, как геометрические свойства нейронов решетки и активность нейронов места преображаются, когда животное обучается ассоциировать определенные точки пространства с наградой. Паттерн активности нейронов решетки больше не похож на идеальную гексагональную решетку.
Группа из Стэндфорда тестировала активность нейронов решетки у крыс в двух условиях (рис. 3): при свободном исследовании местности и при навигации, задействующей пространственную память [8]. В первой арене размером 1,5 × 1,5 метра еда была разбросана случайно, и животные свободно бродили, подбирая ее. Во второй, такого же размера, крыс учили ассоциировать небольшую заплатку пространства со звуковым сигналом и вознаграждением, и по сигналу крысы направлялись в выученную, никак не помеченную область.

Рисунок 3. Схема эксперимента по ориентации в пространстве с наградами и без. а — Слева изображена пустая арена, где крысы бродили случайно, справа — арена с вознаграждением, место которого животные запоминали (красный квадрат). б — Траектория движения крысы в пустой арене (слева) и в арене с вознаграждением (справа), в которой по звуковому сигналу крыса направлялась к спрятанной еде, отмеченной красным квадратом.
Паттерн активности нейронов решетки поменялся во второй арене, хотя гексагональная форма и общий размер остались теми же. Во-первых, поменялась ориентация полей (представьте, как соты вращаются по кругу). Во-вторых, поля активности внутри решетки стали ближе друг к другу, хотя размер каждого отдельного поля остался тем же. В-третьих, при навигации во второй арене у нейронов решетки возросла частота потенциалов действия, причем она возросла у полей, находящихся к награде ближе всего (рис. 4). Другие типы клеток, найденные в энторинальной коре, такие как нейроны направления (head direction cells), нейроны границы (border cells) и другие, также поменяли паттерны активности при навигации к заученной цели. Например, нейроны направления повернули вектор в соответствии с направлением вращения полей нейронов решетки.

Рисунок 4. Изменение свойств нейронов решетки при нахождении награды поблизости. а — Активность одного нейрона решетки в пустой арене (ENV1) и в арене с наградой (ENV2). Поля поменяли ориентацию (повернулись по часовой стрелке); цвет полей обозначает частоту потенциалов действия нейрона (желтый — выше частота). б — Увеличение частоты потенциалов действия в зависимости от расстояния до награды.
Группа из Австрийского института науки и технологий тоже тестировала предположение о том, что роль нейронов гиппокампа не ограничивается чисто геометрической разметкой пространства [9]. Они использовали круглую платформу с большим количеством лунок, в трех из которых прятали сыр. Каждый день крысы заучивали новое расположение трех лунок с угощением, пока ученые следили за активностью нейронов места и решетки (рис. 5).

Рисунок 5. Схема эксперимента группы из Австрии. а — Экспериментальная арена. На первом этапе (pre-probe) крысы бродили по пустой арене случайно, на втором (learning) — выучивали три лунки с сыром, на третьем (post-probe) снова бродили по пустой арене. Между этапами крысы отдыхали. б — Примеры активности клеток места (слева) и клеток решетки (справа); посередине показан гиппокамп и его прилегающие области, такие как энторинальная кора.
Пока крысы заучивали расположение сыра, поле активности нейрона места передвигалось ближе к одной из наград, и у нейронов решетки хотя бы одно поле смещалось к награде (рис. 6), причем по мере обучения клеток со смещающимися полями становилось все больше. Этот результат совпадает с выводами группы из Стэндфордского университета. Исследователей также интересовало, насколько долговечны эти изменения, и оказалось, что нейроны решетки сохраняют искривленную топологию до следующего дня, в то время как нейроны места более забывчивы. Но как происходит трансформация одной топологии решетки в другую с течением времени? В процессе обучения поля решетки могут плавно передвигаться к награде или резко перескакивать от одного положения к другому. Подтвердилась вторая гипотеза, которую авторы назвали «мерцанием» (flickering) — быстрым переключением между двумя позициями без промежуточных стадий.

Рисунок 6. Иллюстрация того, как некоторые поля нейрона решетки перемещаются ближе к цели — спрятанному угощению, — и таким образом деформируют решетку.
Две статьи убедительно подтверждают, что топология и свойства гексагональной решетки вовсе не фиксированные, как у GPS-навигатора, а гибкие и адаптирующиеся под окружающую среду и находящиеся в ней стимулы. Вместо ригидной решетки представьте себе граф (вершины, связанные ребрами), в котором вершины имеют разный вес (чем выше частота потенциалов действия, тем больше вес), а ребра, соединяющие узлы — разную длину. Вокруг желаемого стимула (угощения или цели) пространственное разрешение кодирования увеличивается — решетка как бы стягивается вокруг цели (рис. 6), в то время как в однородных и скучных местах разрешение кодирования наоборот уменьшается — мозгу просто нечего приметить в этом месте.
В действительности можно пойти еще дальше: ученые уже начали строить предположения о том, что пространственный код нейронов места и решетки может организовывать не только физическое пространство, но и когнитивное, размещая на гексагональной решетке не только кухню и диван, но и концепции, мысли и непространственные параметры окружающего мира [10]. Например, мы легко используем в обиходе фразы «близкий друг» или «дальний родственник» — пространственные определения для непространственных категорий. Возможно, что это не просто фигура речи, но и способ, с помощью которого гиппокамп легко запоминает эту информацию и позволяет нам ориентироваться в социальной структуре общества. К примеру, если вы знакомитесь с новым человеком, то его самого могут представлять нейроны места, в то время как его отношение к вам и место в вашем социальном окружении будет закодировано нейронами решетки.
В то время как подобные предположения все еще остаются неподтвержденными, ученые убедительно показали, что позиция взгляда при рассматривании визуальных изображений кодируется нейронами решетки [11]. В эксперименте на обезьянах исследователи записывали активность клеток энторинальной коры, пока животные свободно рассматривали изображения. Были найдены клетки, которые активны, только когда взгляд обезьяны находился в узлах гексагональной решетки (рис. 7). Более того, некоторые клетки были активны только тогда, когда обезьяна рассматривала края изображения (функциональный аналог клеток границы) или синхронизировались с определенным направлением саккад (быстрых движений глаз), что аналогично функции нейронов направления. Таким образом, кодирование ориентации в визуальном пространстве очень похоже на кодирование физического пространства.

Рисунок 7. Движения глаз при свободном рассматривании изображений тоже кодируются нейронами решетки. а — Изображения, которые рассматривали обезьяны, и траектории движения из взгляда. б — Траектория движения взгляда обезьяны по одному изображению (серый) и потенциалы действия нейрона в энторинальной коре (красные точки). в — Карта активности нейрона решетки при рассмотрении одного из изображений.
Можно предположить, что нейроны и связи в гиппокампе и прилегающих областях эволюционировали для кодирования физического пространства, но позже были адаптированы для репрезентации непространственной информации, такой как цели, мотивации, воспоминания, знания, идеи и, возможно, многое другое. Так что когда вы вспоминаете информацию, выученную к экзамену, вы буквально осуществляете навигацию в пространстве — в когнитивном пространстве, вероятно функционирующем по тем же законам, что и физическая навигация.
Литература
- GPS в нашем мозге: Нобелевская премия по физиологии или медицине 2014;
- J. O'Keefe, J. Dostrovsky. (1971). The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Research. 34, 171-175;
- Torkel Hafting, Marianne Fyhn, Sturla Molden, May-Britt Moser, Edvard I. Moser. (2005). Microstructure of a spatial map in the entorhinal cortex. Nature. 436, 801-806;
- Maya Geva-Sagiv, Liora Las, Yossi Yovel, Nachum Ulanovsky. (2015). Spatial cognition in bats and rats: from sensory acquisition to multiscale maps and navigation. Nat Rev Neurosci. 16, 94-108;
- David B. Omer, Shir R. Maimon, Liora Las, Nachum Ulanovsky. (2018). Social place-cells in the bat hippocampus. Science. 359, 218-224;
- Yovel Y. and Ulanovsky N. Bat navigation. In: Learning and memory: a comprehensive reference (2nd Edition) / ed. by Byrne J.H. Amsterdam: Academic Press, 2017. P. 333–345;
- Alison Abbott. (2018). 100 bats and a long, dark tunnel: one neuroscientist’s quest to unlock the secrets of 3D navigation. Nature. 559, 165-168;
- William N. Butler, Kiah Hardcastle, Lisa M. Giocomo. (2019). Remembered reward locations restructure entorhinal spatial maps. Science. 363, 1447-1452;
- Charlotte N. Boccara, Michele Nardin, Federico Stella, Joseph O’Neill, Jozsef Csicsvari. (2019). The entorhinal cognitive map is attracted to goals. Science. 363, 1443-1447;
- Jacob L. S. Bellmund, Peter Gärdenfors, Edvard I. Moser, Christian F. Doeller. (2018). Navigating cognition: Spatial codes for human thinking. Science. 362, eaat6766;
- Nathaniel J. Killian, Michael J. Jutras, Elizabeth A. Buffalo. (2012). A map of visual space in the primate entorhinal cortex. Nature. 491, 761-764.



Комментарии
0Чтобы оставить комментарий, необходимо
войти