МикроРНК — чем дальше в лес, тем больше дров
07 февраля 2008
МикроРНК — чем дальше в лес, тем больше дров
- 3080
- 0
- 5
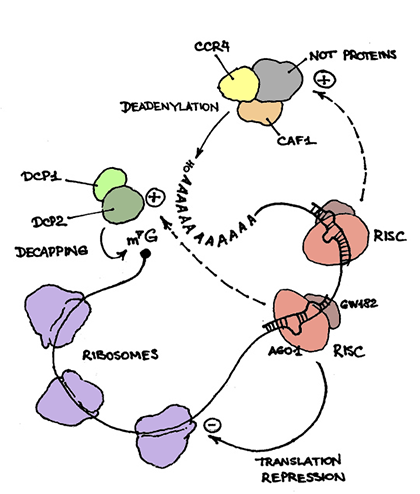
Несмотря на объединённые усилия множества блестящих лабораторий мира, механизм подавления трансляции с участием микроРНК до сих пор во многом остаётся неясным. Более того, природа постоянно вносит свои поправки в наши представления о том, как короткие РНК регулируют экспрессию генов-мишеней.
рисунок Павла Натальина
-
Авторы
-
Редакторы
Традиционной ролью семейства малых интерферирующих РНК (миРНК), — небольших молекул РНК, не кодирующих белков, — считается репрессия генной активности и в частности — синтеза белкá. Однако новое исследование показало (в который уже раз!), что функции одной из групп этих молекул — микроРНК — значительно шире: в определённых случаях они могут стимулировать трансляцию, а не блокировать её.
В последние десять-пятнадцать лет основная догма молекулярной биологии («ДНК → РНК → белок») значительно расшаталась расширилась из-за того, что было открыто великое множество молекулярных механизмов, выбивающихся из этой стройной концепции. Революционными стали открытия, связанные с короткими, не кодирующими белков, молекулами РНК: сюда относятся знаменитое явление РНК-интерференции (отмеченное Нобелевской премией вскоре после открытия) и другие механизмы РНК-зависимой репрессии генов. Одна из разновидностей коротких РНК — микро-РНК (миРНК; miRNA) активно участвует в процессах индивидуального развития организма, включая временной контроль, смерть, пролиферацию и дифференцировку клеток, эмбриональную закладку органов. Они осуществляют «тонкую настройку» экспрессии генов на пост-транскрипционном уровне, тем самым добавляя ещё один уровень сложности в изощрённый механизм внутриклеточной регуляции. Первоначально открытые в «лабораторной» нематоде C. elegans, миРНК были затем обнаружены во многих растениях и животных, а сравнительно недавно и в одноклеточных организмах [1].
Ранее считалось, что короткие РНК используются клетками в процессе РНК-интерференции для специфичной деградации не нужных или вредных РНК [2] — в частности, так клетка может уничтожать чужеродный генетический материал вирусов, родственных им ретротранспозонов и других мобильных элементов, а также РНК, образующихся в результате транскрипции геномных повторяющихся последовательностей. Поэтому логично было предположить, что короткие РНК служат своеобразным прототипом «иммунной системы» внутри клетки. По мере развития наших представлений об участниках и механизмах РНК-зависимой репрессии генов, обнаруживались всё новые и новые интересные особенности, раскрывалось богатое разнообразие существующих в природе путей реализации этой репрессии.
Механизм действия большинства миРНК во многом схож с РНК-интерференцией — короткая (21–25 оснований) одноцепочечная РНК в составе белкового комплекса (ключевой компонент которого — белок семейства Argonaute) с высокой специфичностью связываются с комплементарным участком в 3′-нетранслируемой области (3′-НТО) мРНК-мишени. У растений, миРНК которых полностью комплементарны отрезку мРНК-мишени, связывание приводит к расщеплению мРНК белком Argonaute прямо в середине миРНК–мРНК дуплекса — ситуация, наиболее близкая к «классической» РНК-интерференции. У животных же миРНК комплементарны своей мишени не полностью, и результат связывания получается другим. Долгое время считалось, что связывание приводит к подавлению трансляции (механизм чего до сих пор остаётся загадкой) и не вызывает сколько-нибудь заметной деградации мРНК-мишени. Однако позже было убедительно продемонстрировано, что для большинства миРНК это не так — белки, образующие комплекс с миРНК, стимулируют деградацию мРНК-мишени, привлекая ферменты, осуществляющие удаление кэпа на 5′-конце и укорочение поли(А)-хвоста на 3′-конце мРНК. (С этого обычно и начинается деградация мРНК, отслуживших своё.) Удивительно, но до сих пор не совсем ясно, является ли подавление трансляции причиной или следствием начала деградации мРНК.
Тем временем, жизнь вновь демонстрирует своё нежелание укладываться в какие-либо однозначные схемы: в лаборатории Джоан Стайц (Joan Steitz) было обнаружено, что миРНК могут эффективно подавлять трансляцию, связываясь не только с 3′-нетранслируемой областью мРНК, но и с 5′-НТО [3]. А недавно в журнале Science [4] появилась другая статья этой успешной лаборатории. В ней говорится, что при определенных условиях (напоминающих впадение клеток в «зимнюю спячку», при культивировании их в отсутствие сыворотки в питательной среде) взаимодействие миРНК и мРНК-мишени приводит к строго обратному эффекту — усилению синтеза целевого белка. Это было показано для мРНК одного из цитокинов, фактора некроза опухолей α (TNF-α), и миРНК miR369-3, а потом подтверждено и для миРНК let7-a и miRcxcr4 в паре с искусственно сконструированными мРНК-мишенями.

Рисунок 1. В активно делящейся клетке миРНК, связавшись с комплементарной последовательностью в 3′-нетранслируемом участке мРНК, ингибирует синтез белка (трансляцию). Однако в покоящейся клетке то же самое событие приводит к прямо противоположному эффекту.
Интересно, что действие одной и той же миРНК зависело от состояния клеток: в делящихся клетках миРНК угнетала трансляцию мРНК, а в покоящихся (временно вышедших из клеточного цикла), наоборот — стимулировала (рис. 1). Любопытно и то, что миРНК действовали в составе комплекса, содержащего белки Argonaute 2 и FXR1 (хотя геном человека кодирует 4 родственных белка семейства Argonaute, и все они в той или иной степени имеют дело с миРНК). Именно этим белкам принадлежит главная роль в механизме наблюдаемого явления, тогда как миРНК выполняют функцию «сменного адаптера», через который осуществляется взаимодействие белков с различными мРНК-мишенями.
Вопрос о механизме действия, так же как и лавина других, более конкретных вопросов, вызванных этой публикацией, пока остаются без ответа. А ведь вспоминается время, когда только было открыто явление РНК-интеференции — как же тогда нам всё было понятно и каким же казалось логичным!.. А сейчас можно только развести руками — чем дальше в лес, тем больше дров.
Литература
- МикроРНК впервые обнаружены в одноклеточном организме;
- Григорович С. (2003). Малые РНК в большой науке. Часть 1. Феномен малых РНК. Scientific.ru;
- J. R. Lytle, T. A. Yario, J. A. Steitz. (2007). Target mRNAs are repressed as efficiently by microRNA-binding sites in the 5' UTR as in the 3' UTR. Proceedings of the National Academy of Sciences. 104, 9667-9672;
- S. Vasudevan, Y. Tong, J. A. Steitz. (2007). Switching from Repression to Activation: MicroRNAs Can Up-Regulate Translation. Science. 318, 1931-1934;
- Nicole Rusk. (2008). When microRNAs activate translation. Nat Methods. 5, 122-123.



Комментарии
0Чтобы оставить комментарий, необходимо
войти