Забыть нельзя запомнить
27 февраля 2023
Забыть нельзя запомнить
- 1527
- 0
- 8
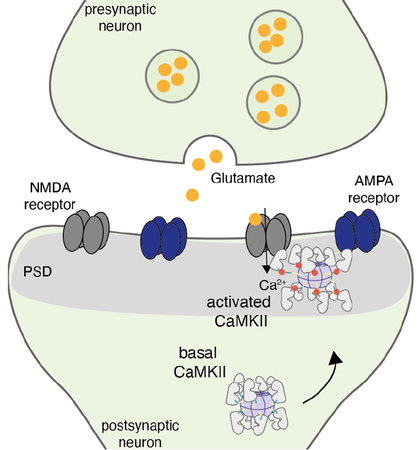
Приток в нейрон Ca2+ приводит перемещению CaMKII к глутаматным рецепторам (AMPA- и NMDA-рецепторам) и соединению с ними, что усиливает передачу нервных импульсов через синапс.
Рисунок в полном размере.
-
Автор
-
Редакторы
Статья на конкурс «Био/Мол/Текст»: Что такое процесс запоминания? Справедливо ли делить память только на долговременную и кратковременную? Как и с помощью чего взаимодействуют эти виды памяти между собой? И, самое главное, какие процессы на молекулярном уровне отвечают за формирование памяти?

Конкурс «Био/Мол/Текст»-2022/2023
Эта работа опубликована в номинации «Школьная» конкурса «Био/Мол/Текст»-2022/2023.
«Книжный» спонсор конкурса — «Альпина нон-фикшн»
Память — это высшая психическая функция, заключающаяся в фиксации, хранении и воспроизведении информации. Уже более века принято делить память на долговременную и кратковременную. Для целей, имеющих значение в течение длительных периодов в жизни индивида, используется долговременная память как хранилище информации, а для целей, актуальных в течение непродолжительных периодов (порядка 20 с), — кратковременная. Источником информации, содержащейся в кратковременной памяти, является сенсорное восприятие и долговременная память. Во время мыслительной деятельности возникает новая информация, которая может использоваться для продолжения интеллектуальной работы и последующего принятия решений, причем эта новая информация может изменяться (корректироваться, заменяться, обновляться), то есть оперативно использоваться в работе по решению текущей задачи. Эта часть памяти получила название оперативной (рабочей).
Как именно формируется память на молекулярном уровне?
В нашей нервной системе есть множество нервных клеток, и между ними имеются различные связи. Связь между нейронами осуществляется за счет синапсов. Типичный синапс состоит из терминали аксона одного нейрона и шипика другого.
Существует несколько моделей памяти; в данной статье мы рассмотрим связь между молекулярными и функциональными механизмами памяти в рамках модели, в которой ведущая роль отводится синапсам (местам соединения между синапсами) и нейронным путям, по которым движутся нервные импульсы. Если мы примем, что память формируется за счет изменения структуры синапсов, их усиления или ослабевания с течением времени в зависимости от активности их работы, то мы можем считать, что память — это результат синаптической пластичности (способности нейронов изменять связи друг с другом внутри нейронных сетей в ответ на их задействование в передаче импульсов). С электрофизиологической точки зрения, синаптическая пластичность — это изменение силы синапса, то есть величины изменения трансмембранного потенциала клетки-мишени в ответ на воздействие определенной силы на пресинаптический нейрон. Пластические изменения происходят в областях мозга, связанных с памятью, и они необходимы и достаточны для хранения этой информации.
Стимуляция и последующее возбуждение нервных клеток лежит в основе всех психических процессов, в том числе и памяти. Поскольку не все нейроны организма заняты в запоминании конкретной информации, необходимо их определенным образом пометить. При слабой стимуляции нейронов активируется специальная метка, нахождение которой в определенных синапсах (задействованных в запоминании данной информации) помечает эти синапсы и нейроны как участвующие в работе по запоминанию конкретной информации. Это (синаптическое мечение) — крайне сложный процесс, в котором задействовано очень много факторов. Покажем работу метки на примере белка CаMKII —кальций/кальмодулин-зависимой протеинкиназы II [1]. Это очень важный фермент, который присутствует в нейронах, концентрируется преимущественно в шипиках и вызывает физиологические процессы, которые значительно увеличивают плотность синапсов. CаMKII настолько распространен, что его доля составляет до 2% от общего белка мозга. CaMKII располагается невдалеке от актиновых филаментов (структурных компонентов клетки), крепко связываясь с ними. В связанном состоянии он неактивен. Для того, чтобы активировать CaMKII, существует несколько способов. Самый эффективный из них — это появление Са2+. Когда это происходит, Са2+ вытесняет CaMKII, сам связываясь с актином. Это заставляет CaMKII перераспределиться внутри шипиков и делает его активным. Активация молекул CaMKII позволяет связываться им с глутаматными рецепторами. Это рецепторы крайне важны для создания памяти. CaMKII, контактируя с внутриклеточными частями глутаматных рецепторов, приводит к их активации, в результате чего происходит усиление синаптических связей. Усиление заключается в увеличении постсинаптического потенциала, возникающего при стимуляции пресинаптических волокон после определенного воздействия (в нашем случае, при воздействии стимула, вызывающего вход Са2+ в нейрон). Иными словами, при повышении концентрации свободного (активного) CaMKII в шипиках облегчается возбуждение постсинаптического звена межнейронных связей, что и лежит в основе синаптической пластичности, которая обеспечивает запоминание.
Метка «живет» недолго. Этот процесс возникновения метки в определенных нейронах (активации CaMKII) связан с молекулярным механизмом кратковременной памяти. Метка помечает синапс как стимулированный. Если вслед за формированием метки сильного стимула не последует, метка распадется. Если же последует сильный стимул, то в клетке произойдет синтез большого количества белков, регулирующих пластичность. Белки, регулирующие пластичность, синтезируются в теле нейронов и стволе дедритов при активации синапсов и транспортируются к определенным дендритным шипикам, где участвуют в усилении синаптической передачи в конкретных синапсах, которая наблюдается при прочном (долговременном) запоминании. Затем метка свяжется с этими белками, и образуется прочная долговременная связь задействованного в запоминании данной информации пула синапсов и синаптических путей в единое целое.
Для создания долговременной памяти обязательно соблюдение следующих условий.
Во-первых, необходим сильный стимул, запускающий синтез белков, регулирующих пластичность. Как описано выше, при слабом сигнале происходит только активация метки. Метка активируется, и белки синтезируются независимо, но для запуска процесса формирования долговременной памяти нужны оба.
Во-вторых, существуют временные ограничения. Необходимо, чтобы и метка, и белки, регулирующие пластичность, синтезировались в одно время. Если белки будет синтезированы после распада (деактивации) метки, то процесс не начнется. Метка существует примерно 30 минут. Белки, регулирующие пластичность, «живут» в течение 1–2 часов. Однако не существует четких рамок — в зависимости от условий протекания реакции, внешних и внутренних факторов возможно как увеличение, так уменьшение окна совпадения.
В-третьих, накладываются и пространственные ограничения. Белки и метка должны находиться в одном месте, причем должна быть возможность контакта между ними.
В итоге минимальными условиями для формирования долговременной памяти является активация метки в ответ на стимул, а также создание временных и пространственных условий, при которых белки, регулирующие пластичность, окажутся в одном месте в одно время с меткой. Иными словами, в каждом синапсе совпадение метки с синтезом белка приводит к поддержанию изменений, вызванных синаптической пластичностью.
Таким образом, формирование долговременной памяти вызывается активацией некоторых специфических контактов нейронных клеток в общей сети. В свою очередь, эта активация может установить метку, способную определять место, где белки, регулирующие пластичность, следует использовать, и цели, для которых их следует использовать. Цель этих белков — установить долговременное облегчение передачи импульсов через синапсы в определенных, участвующих в запоминании конкретной информации, нейронах. Стимулы, направленные на формирование усиления связей между этими нейронами, имеют двойную роль: запускают установку молекулярной метки обучения и синтез белков, регулирующих пластичность. Таким образом, молекулярный механизм долговременной памяти реализуется через одновременную работу более, чем одного элемента — меток и белков, регулирующих пластичность.
Сейчас изучение работы мозга является одним из самых перспективных направлений в науке. Однако, несмотря на все старания ученых, это направление продвигается медленно. Возможно, через несколько лет мы получим больше информации о различных психических процессах, в том числе, и о памяти. Может быть, когда-нибудь люди смогут сами решать, что запомнить, а что не запоминать, смогут мгновенно получать любые знания и «загружать» в память любую информацию. А как по-вашему, какие еще возможности могут предоставить человечеству открытия в механизмах, формирующих память?
Литература
- Nane Griem-Krey, Andrew N. Clarkson, Petrine Wellendorph. (2022). CaMKIIα as a Promising Drug Target for Ischemic Grey Matter. Brain Sciences. 12, 1639;
- Johannes W. Hell. (2014). CaMKII: Claiming Center Stage in Postsynaptic Function and Organization. Neuron. 81, 249-265;
- Ashok Kumar. (2015). NMDA Receptor Function During Senescence: Implication on Cognitive Performance. Front. Neurosci.. 9;
- Diego Moncada, Fabricio Ballarini, Haydée Viola. (2015). Behavioral Tagging: A Translation of the Synaptic Tagging and Capture Hypothesis. Neural Plasticity. 2015, 1-21;
- Dennis Norris. (2017). Short-term memory and long-term memory are still different.. Psychological Bulletin. 143, 992-1009;
- Kosuke Okuda, Kristoffer Højgaard, Lucia Privitera, Gülberk Bayraktar, Tomonori Takeuchi. (2021). Initial memory consolidation and the synaptic tagging and capture hypothesis. Eur J Neurosci. 54, 6826-6849;
- Покровский В. М. Физиология человека. 3-е изд. М.: «Медицина», 2011. — 664 с..



Комментарии
0Чтобы оставить комментарий, необходимо
войти