Эволюция между молотом и наковальней, или Как микробиология спасла эволюцию от поглощения молекулярной биологией
19 января 2010
Эволюция между молотом и наковальней, или Как микробиология спасла эволюцию от поглощения молекулярной биологией
- 8106
- 0
- 14
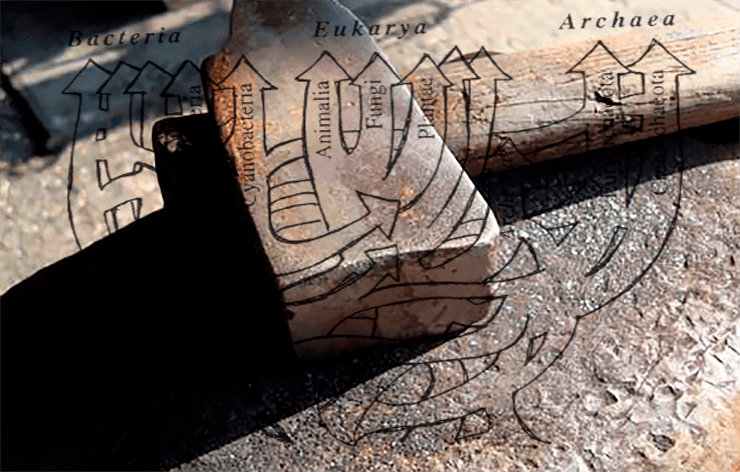
Открытие горизонтального переноса генов стало одним из поворотных моментов в истории молекулярной биологии, отвратив её от погружения в схоластический редукционизм
-
Авторы
-
Редакторы
Заявить в ХХI веке, что молекулярная биология зашла в тупик, — для этого надо иметь не только смелость, но и недюжинный ум. Чтобы опубликовать статью на эту тему в одном из ведущих молекулярных журналов, требуется быть исключительно эрудированным и авторитетным учёным. Мы представляем адаптированный перевод одной из наиболее ярких научных статей прошлого года, написанной двумя выдающимися учёными — Карлом Вёзе и Нигелем Голденфельдом. В работе говорится об опасностях и неудачах современной молекулярной биологии.
Нет границ в свободном поиске. Наука — не место для догм. Учёный имеет право и обязан задавать любые вопросы, ставить под сомнение любые утверждения, искать любые доказательства, исправлять любые ошибки.
Роберт Оппенгеймер. Открытый разум (The open mind. Simon and Schuster, 1955)

Рисунок 1. Карл Вёзе (слева) и Нигель Голденфельд (справа) — авторы статьи. Карл Вёзе (Carl Woese; род. 15 июля 1928) — американский учёный, по образованию физик. Предложил использовать 16S рРНК как универсальную молекулу для классификации организмов. Первым выделил Археи в отдельное царство, отделив их от Эубактерий. Нигель Голденфельд (Nigel Goldenfeld) — американский учёный, по образованию физик, изучает проблему зарождения жизни на Земле.
Nigel Goldenfeld: The humility of ignorance (правое фото)
Эта статья о том, как биология ХХ века фактически пренебрегла изучением наиболее важной проблемы биологии, а именно — эволюции. По сравнению с бурным прогрессом молекулярной биологии, изучение проблем эволюции и по сей день является делом второстепенным. До недавнего времени эволюция была разделом биологии, погрязшим в трясине мелкой схоластики, маскирующей тот факт, что развитие её застыло на уровне достижений прошлого века. Однако, дорогие читатели, не бойтесь: несмотря на такой пессимистичный старт, наш рассказ закончится хорошо. Новейшие данные микробиологии избавили биологию от прокрустова ложа догм, где она находилась так долго.
Такое случалось в науке и раньше. Лучшей иллюстрацией будет удивление, вызванное нашими утверждениями о тупике в биологии, у читателя, убаюканного удивительными достижениями биологической науки XX столетия. О каком тупике речь? Чтобы провести аналогию, мы сошлёмся на схожую ситуацию, приключившуюся с физикой прошлого века. 27 апреля 1900 года в Королевском обществе Великобритании лорд Кельвин (Уильям Томсон) выступил с докладом [1], который должен был подвести итоги работы физиков по созданию единой теории мира. Тема его доклада была: «Облака XIX века над динамичной теорией тепла и света». Им было упомянуто о многих триумфальных достижениях XIX века, таких как:
- Атомная теория Дальтона, которая объяснила феномен материи, и, сомкнувшись со статистической механикой Максвелла и Больцмана, сформировала понятие тепла как формы энергии, связанной с движением атомов или молекул.
- Закон сохранения энергии и принцип увеличения энтропии (начала термодинамики).
- Открытие феномена электромагнетизма, приведшее к предсказанию радиоволн и окончательно объяснившее природу света.
В этом докладе ощущение завершённости физической картины мира било через край, и учёные уверились в скором создании полного теоретического описания всего сущего, так как все основные законы природы уже казались выведенными. Ньютоновское мировоззрение достигло апофеоза, выглядело завершённым и триумфально перешагнуло из XIX в ХХ век. Кельвин очень метко сказал тогда: «Красота и ясность динамической теории, которая описывает свет и тепло с помощью свойств движения, в настоящее время затмевают только два облачка».
Под такой аналогией Кельвин подразумевал, что физики уже почти завершили создание полной картины мира, и только несколько экспериментальных нестыковок омрачают её, хоть и не портят общей картины. Следовательно, единственная оставшаяся работа для учёных — это навести финальный лоск на здание физики, а заодно «разогнать» эти два туманных сгустка. Работа, конечно, немалая, но уже чисто техническая, не нуждающаяся в грандиозных прозрениях. Тем не менее, лорд Кельвин наверняка отдавал себе отчёт, что «облака» заслуживают самого пристального внимания, и сам приложил руку к тому, чтобы рассеять одно из них. Однако все результаты, полученные с помощью классических методов, позже вынудили его признать — эта ноша оказалась неподъёмной.
Первым «облаком» был эксперимент Майкельсона-Морлея, который не обнаруживал никаких следов эфира, заполнявшего, согласно прежним представлениям, всё сущее пространство. Результаты опыта убедительно опровергли теорию эфира, постулируя инвариантность скорости света, и, в конце концов, привели к появлению специальной теории относительности Эйнштейна. Второе «облако» (которое Кельвин пытался развеять лично) было связано с видимым нарушением статистических механических принципов теплоёмкости в газах — артефакт, который мог быть объяснён только с позиций квантовой теории.
Сегодня мы знаем, что «облака» лорда Кельвина предвещали настоящий ураган, и очистить небо от них на тот момент было попросту невозможно. Вместе эти два облака привели к полной и невероятной трансформации физики, для чего потребовалось почти целое столетие. Облака Кельвина были предвестниками новой физики, появления новых фундаментальных идей, которые потрясли ньютоновские устои. Несмотря на то, что всё это относительно хорошо известно, не так часто подчёркивалось, насколько шокирующими были эти открытия — через столько лет работы, после почти завершённой картины мира, физика снова была отброшена от финишной черты далеко назад. Это были примерно те же ощущения, которые испытывает альпинист после непомерно высокого и крутого подъёма: уставший, но в приподнятом настроении, он вдруг находит себя не на вершине, а только на уступе посередине подъёма. Так это было в ХХ веке физики, во времена лорда Кельвина.
Что же происходит с биологией сегодня?
Несмотря на то, что 2009 год был отмечен не одним триумфальным подтверждением теории эволюции [2], бóльшая часть внимания была отдана личности Дарвина (в связи с его юбилеем [3]) и историческим обстоятельствам, сопровождавшим становление теории естественного отбора. Но самой эволюции в теле молекулярной биологии по-прежнему достаточно мало. Молекулярная биология повторяет ошибки микробиологии, о которых мы сейчас и поведём рассказ, — ну а собственно к молекулярной биологии вернёмся чуть позже.
Мир микробов: потерянный и вновь обретённый
Первые сто лет своего существования микробиология боролась за выживание: во-первых, надо было создать методики и приёмы изучения микроскопического мира, а, во-вторых, накопить экспериментальную базу. С такими проблемами сталкивается любая новая наука. Необходимо было упорядочить и классифицировать новые факты — определить правильные категории, выделить ключевые понятия, ввести свою терминологию — всё то, что служит фундаментом для будущих исследований. В случае микробиологии основные понятия остались неизменными и по сей день. Микробиология была «скитальцем» почти весь ХХ век: всегда — прикладная наука; иногда помощник биохимии, иногда — генетики, иногда — молекулярной биологии. Она служила инструментом для решения проблем этих дисциплин. Метания из стороны в сторону привели к тому, что вклад микробиологии в развитие биологии в целом в прошлом веке был значительно меньше того, каким он мог бы быть.
Мало кто знает сегодня, что в начале ХХ века микробиология могла пойти другим путём. В этом случае она выросла бы в науку, мало схожую с микробиологией сегодняшней. Это привело бы к метаморфозе микробиологии в одну из центральных биологических наук, и она не слилась бы с молекулярными дисциплинами и не стала их «падчерицей». Это было бы возможно, если бы микробиологи следовали идеям Мартинуса Бейеринка — наверное, одного из гениальнейших микробиологов в истории. Во время награждения его медалью Левенгука в 1905 году его попросили описать текущее состояние дел. Он ответил:
...Мой подход может быть определён как изучение микробиологической экологии, то есть связи между условиями окружающей среды и соответствующими им формами жизни. Я убеждён, что наиболее важное и плодотворное направление биологии — это знания о той части природы, к которой относятся мельчайшие представители живого. Это, в свою очередь, постоянно напоминает нам о вопросе происхождения самой жизни...
Сто лет назад Бейеринк очертил сегодняшнюю микробиологию — комплексную дисциплину, которая имеет охват от уровня организма до экологии микробных сообществ и их эволюции. Он не упрощал изучение мира микробов. Он понимал, что никакой организм невозможно постигнуть отдельно от его экологии, от совокупности факторов среды, в которой он находится. Он понимал, что отдельный организм и популяция — разные, несводимые к одному понятия, которые нельзя рассматривать в отрыве друг от друга. И он понимал, что мир микробов является центральным пунктом в вопросе о происхождения жизни, а также яркой иллюстрацией процесса эволюции живой материи.
Хотя из общих соображений вроде бы понятно, что организм, особенно микроскопический, нужно рассматривать исключительно внутри популяции и вида, в реальности в микробиологии доминирующей парадигмой стал редукционистский подход — когда система изучается путём дробления на элементарные части и процессы, что зачастую приводит к потере понимания взаимосвязи между ними. К сожалению, взгляды Бейеринка не пережили его самого. Его ученик Ян Клюйвер (Jan Kluyver) был, возможно, чрезмерно впечатлён успехами биохимии, и завернул всю микробиологию на путь редукционизма, всецело подчинив её биохимии. Микробиология Клюйвера стала всего лишь составной частью биохимии, сделав организм вторичным по отношению к метаболизму. Организм, как яркая упаковка, был обречён на растерзание, чтобы обнажить ценную молекулярную начинку. Кстати, такое представление до сих пор популярно среди ряда подразделов микробиологии.
И вот в середине ХХ века микробиология зашла в тупик, — одной только биохимии перестало хватать для описания принципов существования мира микробов. Ни изучение на уровне организма, ни изучение на уровне популяции не могли стать достаточными при таком редукционистском подходе (это возражение, кстати, справедливо и для других дисциплин, таких как молекулярная биология или генетика).
Об ограниченности биохимического подхода в микробиологии впервые было заявлено Станиером (Stanier R.Y.) в 1962 году, когда он и Ван Нель (van Niel C.B.) заявили, что главные результаты микробиологии являются «полным позорищем», неудачной наукоподобной попыткой познать основополагающие принципы организации живого [4]. Авторы определили результаты как «отсутствие ясных представлений о бактериях» и отметили, что «проблема определения бактерий как отдельной группы, исходя из их организации, до сих пор остаётся нерешённой и чрезвычайно важной задачей».
Авторы предложили решение, выдвинув так называемую прокариотическую гипотезу, по которой все безъядерные одноклеточные организмы выделяются в отдельное царство (Procaryota). По тем временам предположение, что все бактерии имеют схожее клеточное строение, было настоящим прорывом и определило дальнейшее развитие микробиологии во второй половине столетия. Более того, Станиер недвусмысленно связал схожесть клеточной организации с единым происхождением, написав, что «...все эти организмы имеют общие структурные характеристики... следовательно, мы можем предположить общего предка в далёком эволюционном прошлом».
Допущение Станиера и Ван Неля о том, что все бактерии относятся к единой группе прокариот, было в высшей степени необходимым для того времени (хотя и неверным — как известно теперь) [5]. Многие научные лаборатории мобилизовались для проверки этой гипотезы. Один из участников работы (и автор этой статьи) — Карл Вёзе — писал в 1969 году Френсису Крику:
...если мы когда-нибудь распутаем направление хода эволюции прокариотических клеток, следующим нашим шагом должна быть расшифровка ещё более ранних этапов эволюции... надо будет заглянуть назад ещё на миллиард лет, чтобы постигнуть такое событие, как возникновение клеток...
Шаг за шагом в царстве прокариот всё более и более ясно вычленялись две отличные между собой группы. В конце 70-х годов ХХ века молекулярные исследования в микробиологии дали первые однозначные результаты. Действительно, было найдено много фундаментальных отличий между эукариотами и всеми бактериями. Но открылась также большая разница и внутри группы бактерий, которые уже фактически стали синонимом слова «прокариоты». Поэтому царство прокариот трансформировали в два отдельных царства (рис. 2): Архебактерии (Archaea) и Эубактерии (Bacteria), а таксон прокариот при этом получил статус надцарства. Хотя это не заставило микробиологов отказаться от концепции прокариот в целом, «эра прокариот» (т. е. деление всего живого только на два царства — прокариот и эукариот) быстро закончилась, и многие микробиологи (особенно те, что помоложе) не потратили лучших лет на обсуждение самого понятия прокариот, переключившись на более важные задачи.

Рисунок 2. Дерево жизни. Всё живое делят на три царства (иногда называемых доменами) — эукариоты, археи и эубактерии. Последние два до 1970-х годов объединяли в единое царство прокариот.
Описанные выше работы позволили микробиологии, наконец-то, построить прочный фундамент для классификации всего мира микроорганизмов. Освобождённая от противоречий в истоках, микробиология снова воссоединилась с экологией. Появление чёткой классификации микроорганизмов — разделение их на архебактерий и эубактерий в 1970-х годах — по праву можно назвать революцией в микробиологии, которая была сопоставима с революцией Коперника в астрономии. Ведь сейчас мы знаем о том, что астрономы, изучая Вселенную, изучали только 4% всей материи — остальное заключено в загадочных тёмной материи и энергии. Так же и мы сейчас понимаем, что по генетическому многообразию мир микробов не только преобладает в биосфере, но и практически не познаваем до конца (это даже если не брать в расчёт виросферу, которую мы рассматриваем как неотделимый атрибут мира микробов). Все современные открытия не только не смогли разрешить вопросов в создании «универсальной» таксономии, но и поставили другой, принципиально неразрешимый вопрос: а существует ли концепция вида, которую можно применять одновременно ко всем представителям микробного мира?
Как известно, во всех хороших рассказах интрига появляется там, где повествование претерпевает перелом. Итак, чтобы следовать этому канону, сейчас мы резко изменим направление нашего повествования и перейдём к молекулярной биологии.
Молекулярная биология: ещё не всё потеряно
Сложно осознать, что дисциплина, которая определила развитие всей биологии в целом на протяжении всего последнего столетия и которая принесла столько знаний и пользы для современного общества, несёт в своей сердцевине фундаментальный изъян. Но, к сожалению, это так.
Молекулярная биология ошеломляюще быстро завоевала признание в рамках классического «ньютоновского» мировоззрения, вооружившись преимущественно редукционистским образом мысли. Хотя на словах всем очевидно, что, сложив вместе описания отдельных механизмов, автоматически целого не получишь, — всё же большинство открытий сделано именно в этом приближении.
В этом «ньютоновском» мире биология стала как бы производной отраслью физики, по сути — инженерным делом без каких-либо фундаментальных задач. Биология стала наукой о биологических машинах, их частях и взаимодействиях этих частей, упражняясь в описаниях, но не объясняя сути объектов.
Однако предназначение биологии состоит не в доскональном описании объектов как моментальных снимков, а в философско-эволюционном понимании феномена их существования. Биология изучает не то, что есть, а то, что является. Другими словами, мы должны стараться ответить на вопрос — что есть жизнь? В какой момент неживые органические молекулы образуют живую материю? Понять этот качественный скачок в организации природы невозможно, лишь изучая составляющие её детали по отдельности. Для нас прелесть макромолекулярного мира состоит в поразительном факте его существования при всей его невероятной сложности. Сейчас молекулярная биология повторяет путь физической науки на прошлом рубеже веков: когда старая, классическая физика уже не могла подступиться к проблемам нелинейного мира из-за его переменчивости, когда простого описания статического мира частиц с линейными связями между ними стало недостаточно.
Общий вектор развития в этих двух случаях схож, тогда как детали отличны. Так, в начале ХХ века редукционизм молекулярной биологии был довольно безобиден и даже полезен — особенно потому, что такой подход позволял быстро накапливать новые данные. Однако потом молекулярная биология начала упрощать всё здание биологии по своему подобию, — вот когда ущербность редукционистского подхода стала проявлять себя в полной мере. Главной ошибкой в выборе курса биологии было чрезмерное внимание, уделённое проблеме экспрессии гена. Сложность этой задачи отодвинула в сторону остальные фундаментальные вопросы.
А началось всё с изучения феномена биологической специфичности (например, молекулярного «узнавания» фермент-субстрат, антиген-антитело и др). Что определяет эту специфичность — простая смесь несвязанных фенотипических признаков, или же она порождается эволюцией живой материи и суть её следствие? Молекулярные биологи предпочли первую формулировку, чрезмерно сосредоточившись на изучении отдельных молекулярных механизмов, — например, принципов действия антител или ферментов. Изучение специфического молекулярного распознавания в редукционистском ключе породило представление о том, что проблема специфичности целиком сосредоточена «здесь и сейчас», а эволюция тут как будто и ни при чём.
В 1940 году Полинг и Дельбрюк обобщили идею специфичности взаимодействий биомолекул [6], расширив концепцию «молекулярных карманов» (лёгшую в основу популярных метафор биохимии того времени — «ключ–замок» и «рука–перчатка») до представления о комплементарности молекулярных поверхностей. Эта концепция стала общепринятой в биологии, особенно после открытия Уотсоном и Криком комплементарности нуклеиновых кислот, и в рамках неё была сформулирована проблема гена — как основной вектор развития была объявлена расшифровка его структуры, механизмов репликации и экспрессии.
Окончательное признание ключевой роли межмолекулярных взаимодействий типа «ключ–замок» пришло с открытием двойной спирали ДНК Уотсона-Крика [7], — структуры, которая наглядно раскрыла феномен репликации гена. Роль гена в хранении наследственности была красиво объяснена в редукционистских терминах молекулярной биологии — неприкосновенность генетического материала обеспечивается идентичным копированием обеих цепей ДНК. Суть репликации была представлена в виде стереохимических взаимодействий, и картина вырисовывалась такой простой и понятной, что сомнений в верности направления исследований не возникало в принципе. Аналогичным образом вскоре была расшифрована и описана транскрипция генов.
Следующей вехой стала проблема трансляции — считывания генетического кода. После выхода статьи Уотсона и Крика направление поисков стало вроде бы понятным. Например, русский физик Георгий Гамов считал, что бороздки в двойной спирали ДНК могут служить матрицей для связывания аминокислот и последующего создания условий для их полимеризации [8]. Гамов — выдающийся физик, чьи работы касались и изучения космического излучения, и теории большого взрыва, — имел косвенное отношение к биологии, просто его заинтересовала сама проблема. Действительно, после открытия триплетности генетического кода простые расчёты показали отсутствие однозначной математической закономерности в кодировании белков — 64 возможных кодона кодируют только 20 аминокислот. Именно это теоретическое несоответствие в дальнейшем станет поворотной точкой в биологии ХХ века.
Френсис Крик предположил, что нуклеиновые кислоты не могут распознавать аминокислоты или что-либо ещё, что не является нуклеиновой кислотой [9]. Хотя он, конечно же, имел в виду ДНК, и этим заявлением фактически отверг возможность существования РНК-посредников, — но также тем самым Крик почти устранил механистические взгляды, господствовавшие в новейшей биологии, приподняв её выше простой алгебраической суммы физико-химических процессов. Ему оставалось только добавить в эту смесь толику эволюции, чтобы спасти всю биологию от редукционизма. Однако Крик и здесь проявил чудеса изобретательности, найдя способ логично вернуться в лоно принципов «шаблонизации», введя молекулы-посредники между ДНК и аминокислотами — в виде «адаптерной гипотезы» [9]. Вместо прямого узнавания генетического кода аминокислотами (по Гамову) было предложено, что каждая аминокислота крепится к специфическому адаптеру, который как раз служит для распознавания кода. Сегодня хорошо известно, что роль криковских «адаптеров» играют молекулы тРНК, правда, Крик считал, что длина адаптеров должна быть всего несколько нуклеотидов, — а на самом деле размер тРНК значительно больше.
Суммируя вышесказанное, давайте отметим различия между чисто шаблонными взглядами на процессы по принципу «ключ–замок» (в центре которых находятся узнавание и спаривание оснований нуклеиновых кислот) и эволюционной парадигмой, учитывающей всё разнообразие процессов и их развитие во времени. Разберём различия на примере «считывающей» и белоксинтезирующей машинерии (мРНК, тРНК, рибосомы и т. п.). В первом случае (модель «ключ–замок») стереохимическое связывание поверхностей вполне объясняет механизм процесса. С этой точки зрения в самом процессе (т. е. «здесь и сейчас») эволюция не играет особой роли, и ею можно пренебречь. Мы и так сможем понять механизм, просто измерив скорость трансляции в текущий момент. Однако если рассматривать тот же процесс с эволюционной точки зрения, — то есть, дополнить принцип работы данными о развитии этой системы или её аналогов в разных организмах, — наше понимание эволюции трансляционной машинерии станет центральной проблемой биологии и потребует эволюционного описания на уровне всей системы. Эволюция отдельных частей систем укажет, какие узлы являются критичными, а какие — вспомогательными и характерными только для данного вида; отслеживание развития и эволюции молекул подскажут их вероятные функции и будущее всей системы. Картина, которую мы получим, будет отличаться от первой, как трёхмерное полноцветное изображение от чёрно-белой фотографии начала прошлого века.
Почти никто не отметил метаморфозу концепции гена со временем. Реальные проблемы генетики не сводятся только к расшифровке механизма трансляции. Однако сегодня почти все усилия направлены на описание потоков информации, извлекаемой сложными белковыми комплексами из матрицы ДНК и сохраняемой в новой форме — в виде белковых молекул. Дальше на первый план выходят взаимодействие биологических молекул между собой и формирование новых, более сложных уровней информационной и биологической иерархии — от молекул к их комплексам, от комплексов — к органеллам, в итоге образуя то, что мы называем живой клеткой. Молекулярная биология не признаёт переменчивости и принципиальной непознаваемости природы. Генотип уже не связывают с фенотипом — он самодостаточен, и изучается подобно «вещи в себе». С этих позиций трансляционный аппарат представляется не более чем одной из вспомогательных лент общего конвейера, которая ожидает в стороне, пока её не изучат и не опишут (не поняв при этом главного).
Такой путь неожиданно оборачивается тупиком. И тупик приключится ещё до того, как молекулярная биология достигнет заявленных целей, — понять, в какой момент неживое вдруг становится живым.
Эволюция: пока только в зародыше
В окончившемся веке эволюция была ахиллесовой пятой наук о жизни. Было видно, что молекулярная биология доминирует во всём и не считается с эволюцией как с центральной осью живого. В рамках своих теорий, молекулярная биология рассматривала эволюцию просто как занятный исторический прецедент, не признавая решающей её роли в фундаментальном познании живых систем (речь идёт о 80-х и 90-х годах — П. С.).
С другой стороны, в ХХ веке и микробиология не исходила из эволюционных концепций. Действительно, почти всё время дисциплина даже не имела правильного представления о самой себе! Микробиология определялась теми практическими сферами, проблемы которых решала (медицины, промышленности), или служила инструментом для развития других дисциплин, — таких как биохимия. До самого последнего времени эволюции в микробиологии не уделяли существенного внимания.
Базового понимания эволюции как непрерывного процесса не даётся и в процессе обучения. Более чем устаревшие объяснения «естественного отбора» не меняются и не оспариваются с начала прошлого века! Может ли это быть объяснено тем, что с тех пор не появилось никаких новых данных? Вряд ли. Дело в том, что основное внимание современной науки направлено в другие стороны. Старая концепция (несколько освежённая синтетической теорией эволюции) всех устраивала, и не было смысла копаться в прошлом. Это значительно ослабило позиции теории эволюции по сравнению с прорывами в других областях, таких как молекулярная биология.
Что мы имеем сегодня? Всё прошлое столетие мы изучали биологию в отрыве от эволюции, без понимания её истинного значения. Наше понимание организации живого, далёкое даже от приблизительной «точки сборки», всё ещё является слишком механистическим и наивным. Микробиология зашла в тупик в поисках «универсальной» таксономии; молекулярная биология в тупике, сама не осознавая этого — её отчаянные попытки понять ген до сих пор находятся где-то на стадии изучения механизмов экспрессии. Чтобы выйти из тупика, мы должны объединить их с эволюционной теорией, а не сосредотачиваться на узких редукционистских моделях. В двадцатом веке учёные не смогли осознать, что биологическая организация на самом деле — эволюционная проблема.
Эволюция — путь наверх
Конфликт молекулярной биологии и эволюции не может длиться долго. Ведь даже центральный метод всей молекулярной биологии — секвенирование макромолекул по Сенгеру — является также эффективнейшим инструментом эволюционных исследований. Первым на этот потенциал обратил внимание ещё Френсис Крик — основоположник молекулярного редукционизма. Он писал:
...Биологам следует понять, что мы должны создать «белковую таксономию», — то есть, изучить аминокислотный состав гомологичных белков разных видов... необъятный массив эволюционной информации может скрываться в этих данных...
Осознавал ли он сам важность своих слов? Мы не знаем. Однако впоследствии Крик всё дальше и дальше уходил от эволюционных идей вглубь изучения механизмов экспрессии генов, рассматривая трансляцию просто как одну из занятных молекулярных машин, но не как фундаментальный эволюционный феномен.
Эволюционные идеи Крика переоткрыли в середине 1960-х Цукеркандл и Полинг [10], и на этот раз идея обратила на себя внимание. Классическое филогенетическое древо теперь начало дополняться новыми, молекулярными данными. Сравнительные исследования белков-гомологов, полученных из разных видов, таких как цитохром c или гемоглобин, подтвердили многие прежние данные о биологическом родстве разных организмов.
Однако наилучшим объектом, по которому можно изучать «эволюционное родство», оказалась рРНК — с ней было проще работать, поскольку она более консервативна у разных видов и более стабильна при выделении. Мутации в рРНК накапливаются медленно, и количество этих мутаций может использоваться для измерения эволюционного «расстояния» между организмами. Таким образом, рРНК — это почти идеальный «эволюционный хронометр» (см. также: «Сверим часы» [11]).
Из трёх типов рРНК удобнее всего для анализа оказались 16S и 18S рРНК [12]. Анализ рРНК дал возможность обоснованно разбить всё живое на три группы: эукариоты, архебактерии и эубактерии. Исследования родства среди эубактерий показали наличие 10 эволюционных ветвей, причём в пяти из них обнаружены фотосинтезирующие организмы. Это означает, что разные группы фотосинтезирующих бактерий эволюционно ближе к отдельным ветвям нефотосинтезирующих бактерий, чем друг к другу. Эти открытия заставляют по-новому взглянуть на эволюцию одноклеточных организмов и возникновение фотосинтеза [13].
И это было только началом. Не только эволюция, но и вся биосфера стала прозрачной для исследования новейшими молекулярными инструментами. Норман Пейс [14] стал первым, кто разглядел и экспериментально проработал идею о том, что генетическое секвенирование позволяет избавиться от необходимости работать только с культивируемыми организмами (в первую очередь речь идёт о микроорганизмах, выращиваемых на искусственных питательных средах). Он показал, что мы можем просто «считывать» гены интересующих нас организмов и исследовать их отдельно — тогда и необходимость ловить и культивировать сами организмы отходит в прошлое (рис. 3). По сути, его идея как никогда приблизила современную микробиологию к «идеальной» микробиологии Бейеринка и позволила изучать разнообразие микробиологических биосистем прямо в среде их обитания. (Речь идёт, например, об изучении геномов почвенных микроорганизмов («Почва может иметь собственный геном» [15]), микрофлоры человека («Кишечная микрофлора превращает человека в „сверхорганизм“» [16]), о «переписи» морской жизни (Census of Marine Life) и т. п.) Причём вместо того, чтобы клонировать неизвестные гены в «рабочие» бактерии типа Escherichia coli, а затем изучать их, можно провести тотальное параллельное прочтение ДНК всех микроорганизмов, встречающихся в той или иной экологической нише. Технологии секвенирования ДНК нового поколения позволяют создавать библиотеки всех имеющихся фрагментов ДНК и секвенировать такие библиотеки с огромной производительностью. Даже не зная, какому именно виду бактерий они принадлежат, эти данные при посредстве современных биоинформационных технологий могут быть составлены в единую картину.

Рисунок 3. Изучение геномов микроорганизмов из термальных источников позволило обнаружить многие новые виды архей и бактерий, ранее неизвестных по причине их «некультивируемости»
Будущее биологии в эволюции
Пожалуй, ничто так ярко не демонстрирует необоснованность претензии современной синтетической теории эволюции быть всеобъемлющим описанием эволюционного процесса, как ёе первое неудачное столкновение с микробиологией. Впервые это случилось в годы, последовавшие за классическим экспериментом Эйвери, Маклеода и Маккарти на пневмококках, с помощью которого была доказана роль ДНК как наследственного материала. Прямым следствием этой работы было открытие Джошуа Ледербергом (Joshua Lederberg) и коллегами конъюгации у бактерий [17] и последующее открытие трансдукции [18]. Был открыт «неклассический» генетический феномен, который через какое-то время станет несомненным вследствие распространения устойчивости к различным лекарственным препаратам. Однако, в силу влияния синтетической теории эволюции, за редкими исключениями, открытие почти не получило признания в последующие 30 лет, и по-настоящему внимание на горизонтальный перенос генов обратили только в последние несколько десятилетий. Столь продолжительное игнорирование открытия и его важности для понимания механизмов эволюции иллюстрирует догматизм и застой, которые царили не только в синтетической теории эволюции, но и в микробиологии прошлого.
Много лет спустя Ледерберг написал: «...позже я всегда удивлялся: почему подобные простые эксперименты 1946 года не были сделаны вслед за повторным открытием Законов Менделя, в 1900 году. Это позволило бы микробиологии вырваться на 50 лет вперёд...».
Ледерберг был столь заинтригован вопросом, почему его открытие не было сделано много раньше, что детально изучил причины этого «перезрелого открытия». Он объяснил это слишком сильным догматизмом в тогдашней микробиологии. Он писал: «...поскольку было признано, что половое размножение не характерно для бактерий, никто и не предполагал принципиальную возможность передачи ДНК между [бактериальными] клетками...» [19].
Догматическое мышление преобладает слишком часто, оказывая с пагубное влияние на все области микробиологии, молекулярной биологии и эволюции. Например, это привело к некорректному с научной точки зрения представлению о распространении геномной информации у прокариот.
Сегодня мы знаем, что горизонтальный перенос генов — мощная эволюционная сила мира микробов, экологическая суть которой осознана лишь недавно. Долгое время мы удивлялись — почему эукариоты лишены такого мощного инструмента. Однако оказалось, что это очередная недооценка сил эволюции в природе. На сегодняшний день горизонтальный перенос генов достоверно показан у и многих высших эукариот — не только растений, но и у животных, и у человека [20].
Но давайте закончим эту историю на оптимистичной ноте. Мы ещё успеем всё переосознать! Сейчас в биологии наступили захватывающие времена. Геномные технологии создают беспрецедентный инструментарий для познания основ живого. Эволюционные исследования позволяют отбросить столетние догмы и создать новую истинную науку, созданную из сплетённых микробиологии и молекулярной биологии. И если биология XXI столетия не уступит своего места нанотехнологиям, она должна сосредоточиться на познании биологической организации на базе эволюционных взглядов. Биологическая организация теперь уже не должна будет восприниматься иначе как проявление эволюционного процесса. Только таким образом биология станет чем-то большим, чем просто разрозненной коллекцией фактов.
По материалам оригинальной статьи Вёзе и Голденфельда [21]. Авторы «Биомолекулы» не обязательно во всех деталях разделяют точку зрения авторов статьи.
Литература
- Thomson W. (Lord Kelvin) (1901). 19th century clouds over the dynamical theory of heat and light. Philosophical Magazine and Journal of Science. 2, 1–39;
- Рубцов А. (2009). Чарльз Дарвин и теория эволюции. «Наука и жизнь». 1;
- Элементы: «Дарвину 200 лет»;
- Stanier R.Y. and van Niel C.B. (1962). The concept of a bacterium. Arch. Mikrobiol. 42, 17–35;
- J. Sapp. (2005). The Prokaryote-Eukaryote Dichotomy: Meanings and Mythology. Microbiology and Molecular Biology Reviews. 69, 292-305;
- L. PAULING, M. DELBRUCK. (1940). THE NATURE OF THE INTERMOLECULAR FORCES OPERATIVE IN BIOLOGICAL PROCESSES. Science. 92, 77-79;
- J. D. WATSON, F. H. C. CRICK. (1953). Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid. Nature. 171, 737-738;
- G. GAMOW. (1954). Possible Relation between Deoxyribonucleic Acid and Protein Structures. Nature. 173, 318-318;
- Crick F.H.C. (1958). On protein synthesis. Symp. Soc. Exp. Biol. 12, 138–163;
- Emile Zuckerkandl, Linus Pauling. (1965). Molecules as documents of evolutionary history. Journal of Theoretical Biology. 8, 357-366;
- Сверим часы;
- C. R. Woese, G. E. Fox. (1977). Phylogenetic structure of the prokaryotic domain: The primary kingdoms. Proceedings of the National Academy of Sciences. 74, 5088-5090;
- Guttel R., Larsen N., Woese C. (1994). Lessons from an evolving rRNA:16S and 23S rRNA structures from a comparative perspective. Microbiol. Rev. 8, 10–24;
- N. R. Pace. (1997). A Molecular View of Microbial Diversity and the Biosphere. Science. 276, 734-740;
- Войцеховская Я. (2006). Почва может иметь собственный геном. «Полит.ру»;
- Элементы: «Кишечная микрофлора превращает человека в „сверхорганизм“»;
- JOSHUA LEDERBERG, E. L. TATUM. (1946). Gene Recombination in Escherichia Coli. Nature. 158, 558-558;
- Zinder N.D., Lederberg J. (1952). Genetic exchange in salmonella. J. Bacteriol. 64, 679–699;
- Harriet Zuckerman, Joshua Lederberg. (1986). Postmature scientific discovery?. Nature. 324, 629-631;
- Марков А.В. Горизонтальный перенос генов и его роль в эволюции. «Проблемы Эволюции»;
- C. R. Woese, N. Goldenfeld. (2009). How the Microbial World Saved Evolution from the Scylla of Molecular Biology and the Charybdis of the Modern Synthesis. Microbiology and Molecular Biology Reviews. 73, 14-21;
- W. F. Doolittle. (1999). Phylogenetic Classification and the Universal Tree. Science. 284, 2124-2128.



Комментарии
0Чтобы оставить комментарий, необходимо
войти