От сложного к простому: трудности симбиогенеза
17 мая 2016
От сложного к простому: трудности симбиогенеза
- 7594
- 0
- 7
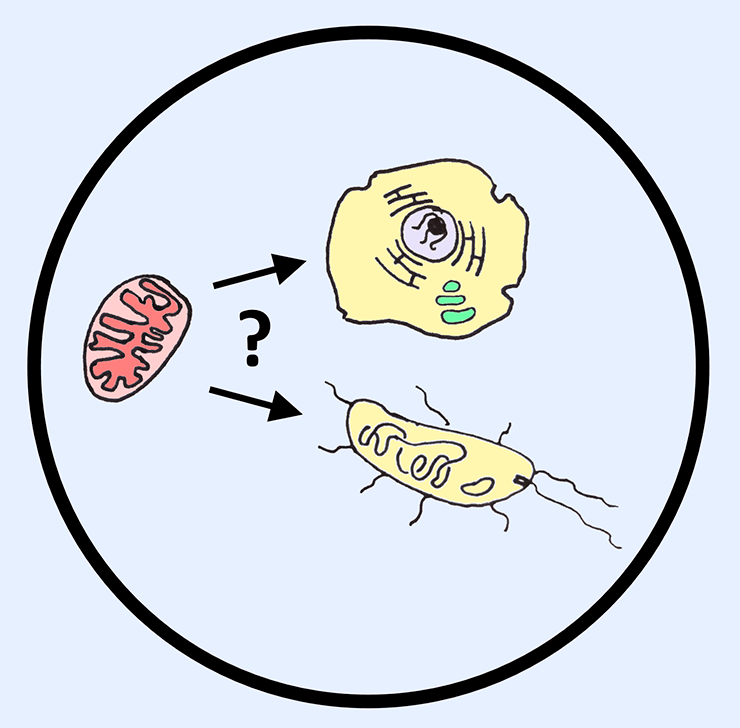
Был ли организм, «приручивший» митохондрию, ядерным? Пока на этот вопрос нельзя ответить абсолютно уверенно.
-
Автор
-
Редакторы
Митохондрии — верные спутники эукариот. Согласно теории симбиогенеза, именно обретение митохондрий спровоцировало формирование ядерных организмов. Одним из доказательств этой теории было обнаружение митохондрий или подобных им органелл у всех, даже самых простых, эукариот. Но в мае 2016 года коллектив чешских ученых описал первый в истории ядерный организм, не содержащий даже косвенных признаков митохондрий. Может ли это открытие пошатнуть современные представления о ранней эволюции эукариот?
Митохондрии — двумембранные органеллы, снабжающие энергией клетки практически всех эукариот. Достоверно известно, что они родственны α-протеобактериям и стали частью эукариотических клеток около 1,5 млрд лет назад [1]. О бактериальном происхождение митохондрий свидетельствует наличие двух мембран (внутренней собственной и внешней хозяйской), собственных кольцевой ДНК и трансляционной машины, а также способности независимо делиться. Некоторые даже полагают, что апоптоз — попытка митохондрии убить поглотившего ее эксплуататора.
Тем не менее в научном сообществе до сих пор нет единого мнения относительно роли этого симбиоза в развитии эукариот (рис. 1). Сторонники теории симбиогенеза утверждают, что слияние некой археи с предком митохондрий (бактерией) запустило цепочку событий, приведших к образованию эукариот современного типа. Приверженцы архезойной теории (гипотезы), наоборот, считают, что «приручить» митохондрию мог только уже оформившийся ядерный организм — архезой [2].
Благодаря Карлу Вёзе и Джорджу Фоксу, в 70-х годах прошлого века сравнившим гены 16S рРНК множества живых существ, долго обманывавшие микробиологов своим визуальным сходством две формы доядерных организмов (прокариот) развели окончательно и бесповоротно, да еще и на высшем уровне: эубактерий лишили права на «истинность» (—эу), архебактерий — права носить гордое имя бактерий, зато в новой системе живых организмов им отвели по собственному домену (таксону высшего ранга): «Эволюция между молотом и наковальней, или как микробиология спасла эволюцию от поглощения молекулярной биологией» [3] и «Карл Вёзе (1928–2012)» [4]. Так в 1990 году человеку предложили осознать, что все живые существа филогенетически разбиваются на три домена: Бактерии, Археи и Эукариоты, — причем бактерии отличаются от архей даже больше, чем археи от эукариот, а недавно вообще чуть ли не усомнились в целесообразности разделения последних: «Нашли предков всех эукариот» [5]. Однако предложение почти тридцатилетней давности до сих пор не нашло понимания у многих авторов отечественных учебников биологии. А что? Вдруг опять эти ученые всё поменяют, а им переписывать каждые -цать лет учебники что-ли? — Ред.
В 1928 году знаменитый биолог рубежа XIX-XX веков Эдмунд Вилсон высказался о гипотезе бактериального происхождения митохондрий так: «Подобные идеи чересчур фантастичны, чтобы их можно было обсуждать в приличном биологическом обществе» [6]. Сегодня же подобное отношение сформировалось к архезойной теории, а ключевая роль митохондрий в ранней эволюции эукариот общепризнанна. Открытие первого истинно безмитохондриального простейшего заставляет еще раз задуматься о сильных и слабых сторонах каждой из теорий.

Рисунок 1. Разница между архезойным (слева) и симбиогенетическим (справа) сценариями.
Теория симбиогенеза
Основным доказательством того, что митохондрии — триггер эволюции эукариот, является их вездесущность. Даже у анаэробных эукариот в клетках остаются следы митохондрий (рис. 2). Например, у паразитического простейшего Entamoeba histolytica митохондрии деградировали до митосом, лишенных ДНК или белков дыхательной цепи [7]. Тем не менее в митосомах транспорт белков идет по типично митохондриальному механизму, а ядерный геном E. histolytica кодирует типично митохондриальные гены. На сегодня известно более 1000 эукариот, не содержащих митохондрий, и все они произошли от более развитых форм, обладающих митохондриями [8].

Рисунок 2. Электронные микрографии митохондриеподобных органелл. а — собственно митохондрии из куриного мозжечка; б — гидрогеносомы из анаэробного грибка Neocallimastix patriciarum; в — митосомы из кишечного паразита Entamoeba histolytica. Масштаб 100 нм.
Одно из преимуществ симбиогенетической теории по сравнению с архезойной в том, что она объясняет возникновение ядра и интронную структуру генома. У прокариот широко распространен горизонтальный перенос генов (ГПГ), за счет которого популяции могут быстро обмениваться частями генофонда [9]. ГПГ способствует незащищенности генома — ведь поступающая ДНК в этом случае ничем не отделена от содержимого хозяйской клетки.
Вполне вероятные попытки клетки-хозяина разрушить пока еще не одомашненного симбионта приводили к высвобождению в цитоплазму симбиотической ДНК. Эта ДНК, будучи в непосредственной близости от хозяйского генома, могла легко в него встраиваться [10]. Из-за ГПГ даже в эукариотах, утративших митохондрии, находят изначально митохондриальные гены.
Такое слияние геномов могло, во-первых, способствовать развитию взаимозависимости симбионта и хозяина. Во-вторых, обильный ГПГ мог переносить не только гены, обеспечившие переплетение метаболизма двух организмов, но и эгоистичные ретроэлементы [11]. Вторжение интронов II группы, вырвавшихся из α-протеобактерий, привело к разрыхлению исходно очень плотного генома хозяина: до 80% хозяйской ДНК теперь были интронами [12], [13]. В такой сложной ситуации клетка-хозяин развила несколько линий защиты своего генома от шквала интронов: возникли система внутренних мембран и ядро, убиквитиновая система деградации поврежденных белков, нонсенс-опосредованный распад РНК и прочие характерные особенности эукариот (рис. 3).

Рисунок 3. Формирование основных черт эукариот можно объяснить последовавшим за митохондриальным симбиозом вторжением интронов II группы в геном клетки-хозяина.
Еще одно мощное доказательство симбиогенеза — энергетические потребности эукариот. Хотя энергопотребление про- и эукариот в пересчете на грамм веса примерно одинаково, ядерные клетки гораздо крупнее безъядерных, из-за чего они потребляют примерно в 5000 раз больше энергии (2300 пВт/кл против 0,5 пВт/кл). При пересчете энергопотребления на средний ген одноклеточного организма оказывается, что эукариотический ген потребляет в 1000 раз больше энергии [8] . Без митохондриальной энергетики было бы невозможно не только создать сложные, большие и активно передвигающиеся организмы, но даже обеспечить функционирование типичных для эукариот клеточных структур.
У гигантских бактерий масштабирование прокариотической энергетики за счет массовой полиплоидизации (как в случае Epulopiscium, дорастающей до 0,6 мм и содержащей 200 000 копий генома размером 3,8 млн п.н. [14]) не приводит к повышению выхода энергии на ген, и клетка остается типично бактериальной [8]. — Авт.
Еще один важный факт, подкрепляющий симбиогенетический сценарий — существование внутриклеточных симбионтов бактерий. Случаи эндосимбиоза у бактерий крайне редки в природе, но всё же они есть и демонстрируют, как мог зарождаться эукариотический домен жизни [15].
Архезойная теория
Архезои — предполагаемые безмитохондриальные, но ядерные предки современных эукариот. Согласно архезойному сценарию митохондрии были одомашнены только на поздних стадиях эволюции эукариот и не оказали на этот процесс значительного влияния.
Одно из основных положений симбиогенеза — гипотеза исходной простоты. О жизни во времена протерозоя известно крайне мало, поэтому о ее устройстве существует множество часто взаимоисключающих предположений. Если по первой гипотезе считается, что от прокариот с очень компактными геномами произошли более сложные эукариоты, то в архезойном сценарии изначально существовали клетки с запутанными и громоздкими геномами, от которых путем редукции произошли более простые прокариоты. Эукариоты же лишь сохранили первичную сложность.
Эволюция геномов и правда далеко не всегда движется от простого к сложному. И среди эукариот есть примеры, подтверждающие это.
Типичный эукариотический геном содержит множество интронов, прерывающих кодирующую часть генов, ретроэлементов и некодирующих повторов, а также протяженные межгенные участки, которым одни приписывают важные регуляторные функции, а другие, не церемонясь, называют «мусорной ДНК». В итоге характерная для эукариот плотность генов — 0,012 гена/т.п.н.; у прокариот же геномы гораздо компактней — 1 ген/т.п.н. [16]. У ряда паразитических простейших плотность генов приближается к прокариотической, но такая компактизация у них всегда сопровождается и потерей тысяч генов их свободноживущих предков. В результате геномы паразитических протист обычно содержат меньше 10 000 генов.
Тем не менее редукция генома совсем не обязательно сопровождает его компактизацию. Доказательства этого можно найти как у простейших, так и у многоклеточных форм жизни.
Например, свободноживущая инфузория Paramecium tetraurelia содержит 30000 генов, на каждый из которых приходится в среднем 2 т.п.н. Такая компактность достигается за счет сокращения размера интронов до предельных 25 п.н. и уменьшения межгенных расстояний [17].
У Caenorhabditis elegans плотность генов достигает 0,2 гена/т.п.н. [18], причем 15% генов этого червя организованы в опероны, как у бактерий [19].
Даже у позвоночных могут быть необычно компактные геномы: геном рыбы фугу в восемь раз меньше человеческого в основном за счет низкого содержания повторов (рис. 4) [20].

Рисунок 4. Рыба фугу имеет необычно компактный для позвоночного геном частично благодаря коротким интронам. Вертикальная ось дана в логарифмическом масштабе.

Рисунок 5. Гипотеза изначальной сложности подразумевает, что прокариотические ветви жизни произошли от более сложных форм за счет редукции. Редукционный вектор развития мог быть задан первыми хищными архезоями, угнетающими прочие организмы.
Приведенные примеры показывают, что простота прокариотических геномов может возникать вторично. Если это так, то LUCA — последний общий предок всех современных организмов — мог обладать геномом эукариотического типа.
Гипотезу исходной сложности также подтверждают так называемые гены-сигнатуры («подписи») — эукариотические гены, не имеющие прокариотических гомологов. Вероятнее всего, эти гены содержались в LUCA, но были утеряны бактериями и археями.
К несчастью архезойной теории, список сигнатур значительно поредел с начала XXI века. Среди множества секвенированных с тех пор геномов были найдены их прокариотические гомологи. [21] Таким образом, с каждым годом становится всё больше белков, чье присутствие у эукариот можно объяснить тем, что их гены были принесены архейным либо бактериальным предком в ходе симбиогенеза.
И в то же время открытие прокариотических гомологов белков клеточного движения (актинов, тубулинов и кинезинов) косвенно подтверждает возможность того, что архезои могли активно передвигаться и даже быть первыми хищниками на Земле, способными к фагоцитозу [22]. Возникновение хищников в автотрофно-сапротрофном сообществе колыбели жизни должно было оказать колоссальное влияние на ход эволюции. В самых захватывающих сценариях одни жертвы архезоя приспосабливаются к быстрому делению и росту, а другие — к нишам, куда архезой не способен проникнуть. В итоге гипотетический архезой повел эволюцию своих современников по редуктивному пути с упором на гибкость метаболизма и скорость деления, в ходе чего сформировались известные нам прокариоты (рис. 5) [23].
Но несмотря на то, что архезойная теория имеет некоторые сильные стороны или как минимум наносит колкие удары в сторону симбиогенеза, ей не хватает главного — она не объясняет, как и почему сформировалось ядро.
Уникальная находка
В 1980-х существовало множество претендентов на звание современного архезоя, но в последующие годы у всех них нашли митохондриеподобные органеллы (митосомы и пероксисомы) и гены-маркеры митохондриального прошлого: гены сборки Fe-S-белков, митохондриальных транспортеров и шаперонов, синтетазы кардиолипина. К тому же некоторые белки, синтезируемые в цитоплазме, обладают последовательностями импорта в митохондрии, которые могут сохраняться и в отсутствие самих митохондрий.
С каждым новым «закрытием» потенциального архезоя безмитохондриальный сценарий становления эукариот оказывался всё менее вероятным. И вот в мае 2016 года наконец-то появился новый потенциальный архезой, не содержащий даже следов митохондрий [24].
Речь идет об анаэробной оксимонаде Monocercomonoides sp. PA203, живущей в кишечнике насекомых. Оксимонады лишены митохондрий и не содержат в ядерной ДНК гены митохондриального происхождения. Энергию они получают из гликолиза, идущего в цитоплазме.
Геном Monocercomonoides sp., расшифрованный коллективом чешских ученых, содержит 16629 генов, среди которых нет названных выше маркеров. Поиски митохондриальных гомологов и белков с импортными последовательностями тоже не дали удовлетворительных результатов (рис. 6).
Единственное, что удалось найти — два гена, продукты которых у близкого родственника Monocercomonoides sp. могут содержаться (а могут и не содержаться) в митохондриях, при этом они лишены импортных последовательностей.

Авторы открытия считают, что Monocercomonoides когда-то всё же содержали митохондрии, поскольку у близкородственных родов есть следы митохондрий. Еще остается возможность, что у этих простейших есть пока не обнаруженные митосомы, деградировавшие настолько, что в геноме не осталось каких-либо признаков их присутствия.
Так или иначе, Monocercomonoides sp. — пока уникальный случай истинно безмитохондриального протиста за всю историю биологии. И этот случай доказывает, что эукариоты могут жить не только без митохондрий, но и без их генетического наследства .
В какой последовательности предки эукариот разживались внутриклеточным скарбом и что стало счастливым билетом в эволюционное будущее, можно узнать из статьи «Генеалогия белков свидетельствует о позднем приобретении митохондрий предками эукариот» [25]. — Ред.
Это открытие, конечно, не наносит сокрушительного удара по теории симбиогенеза, но однозначно заставляет задуматься, что есть необходимость и что есть излишество в эукариотах.
Литература
- Как появились митохондрии (рассказ, похожий на сказку);
- Кунин Е.В. Логика случая. М.: Центрполиграф, 2014. — 527 с.;
- Эволюция между молотом и наковальней, или Как микробиология спасла эволюцию от поглощения молекулярной биологией;
- Карл Вёзе (1928–2012);
- Кондратенко Ю. (2015). «Нашли предков всех эукариот». «Кот Шрёдингера». 6;
- van der Giezen M. (2009). Hydrogenosomes and mitosomes: conservation and evolution of functions. J. Eukaryot. Microbiol. 56, 221–231;
- Tovar J., Fischer A., Clark C.G. (1999). The mitosome, a novel organelle related to mitochondria in the amitochondrial parasite Entamoeba histolytica. Mol. Microbiol. 32, 1013–1021;
- Lane N. (2011). Energetics and genetics across the prokaryote-eukaryote divide. Biol. Direct. 6, 35;
- Закинули археи эволюционный невод и вытянули...;
- Как составлялся геном эукариот: эндосимбиоз VS. непрерывный горизонтальный перенос;
- Rogozin I.B., Carmel L., Csuros M., Koonin E.V. (2012). Origin and evolution of spliceosomal introns. Biol. Direct. 7, 11;
- Koonin E.V. (2009). Intron-dominated genomes of early ancestors of eukaryotes. J. Hered. 100, 618–623;
- Сколько сора в нашей ДНК;
- Тысячекратная полиплоидия гигантской бактерии Epulopiscium;
- von Dohlen C.D., Kohler S., Alsop S.T., McManus W.R. (2001). Mealybug beta-proteobacterial endosymbionts contain gamma-proteobacterial symbionts. Nature. 412, 433–436;
- Lane N. and Martin W. (2010). The energetics of genome complexity. Nature. 467, 929–934;
- Zagulski M., Nowak J.K., Le Mouël A., Nowacki M., Migdalski A., Gromadka R. et al. (2004). High coding density on the largest Paramecium tetraurelia somatic chromosome. Curr. Biol. 14, 1397–1404;
- Hillier L.W., Coulson A., Murray J.I., Bao Z., Sulston J.E., Waterston R.H. (2005). Genomics in C. elegans: so many genes, such a little worm. Genome Res. 15, 1651–1660;
- Blumenthal T., Davis P., Garrido-Lecca A. (2015). Operon and non-operon gene clusters in the C. elegans genome. WormBook. 28, 1–20;
- Aparicio S., Chapman J., Stupka E., Putnam N., Chia J.-M., Dehal P. et al. (2002). Whole-genome shotgun assembly and analysis of the genome of Fugu rubripes. Science. 297, 1301–1310;
- Koonin E.V. (2010). The origin and early evolution of eukaryotes in the light of phylogenomics. Genome Biol. 11, 209;
- Mayer F. (2003). Cytoskeletons in prokaryotes. Cell Biol. Int. 27, 429–438;
- Kurland C.G., Collins L.J., Penny D. (2006). Genomics and the irreducible nature of eukaryote cells. Science. 312, 1011–1014;
- Karnkowska A., Vacek V., Zubáčová Z., Treitli S.C., Petrželková R., Eme L. et al. (2016). A eukaryote without a mitochondrial organelle. Curr. Biol. 10, 1274–1284;
- Элементы: «Генеалогия белков свидетельствует о позднем приобретении митохондрий предками эукариот».



Комментарии
0Чтобы оставить комментарий, необходимо
войти