Как обойтись без полового размножения, не отказываясь от его преимуществ?
18 июня 2015
Как обойтись без полового размножения, не отказываясь от его преимуществ?
- 609
- 0
- 2
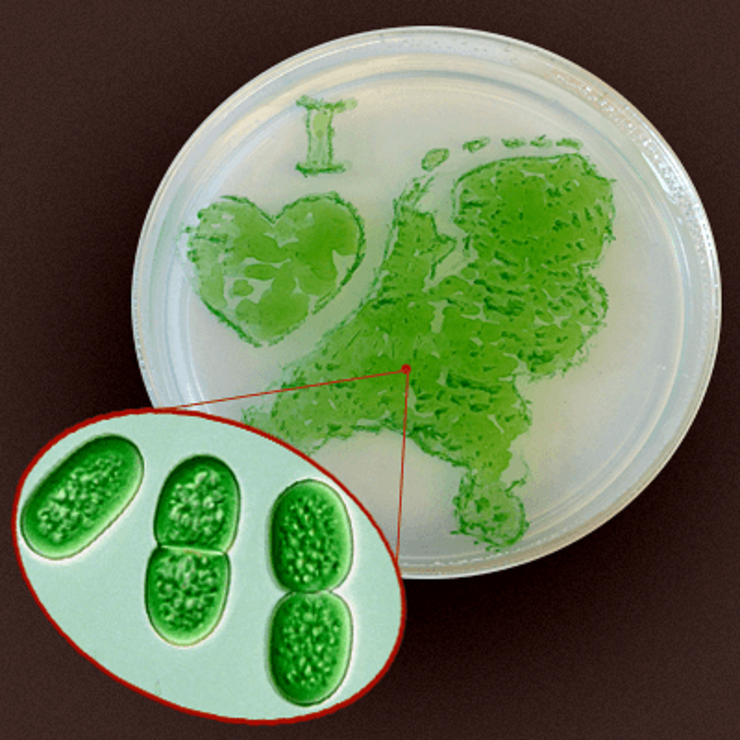
Фотосинтезирующие Нидерланды. Любовь к родине голландские биологи выразили, вырастив цианобактерию Synechococcus elongatus в чашке Петри, содержащей только воду и соли. На свету бактерия строит колонии и формирует биопленки буквально из воздуха — она ведь фотосинтезирующая. И потому, между прочим, рассматривается как кандидат «в биотопливо».
Рисунок в полном размере.
основное фото с сайта sitn.hms.harvard.edu
-
Автор
-
Редакторы
Ваш вид не размножается половым путем, а генетическое разнообразие нужно как-то поддерживать? Если вы — часть биопленки, то выход есть.
Смысл полового размножения в том, чтобы создавать новые комбинации генов. И дело не только в том, что ребенок получает часть генетического материала от папы, а часть — от мамы. Важно, что еще до того, как передать часть своих генов ребенку, наследственный материал каждого из родителей тасуется. В результате они передают по наследству не совсем те хромосомы, что были у них самих. Этот процесс перетасовки генов между копиями своих хромосом называется кроссинговером, и ему мы обязаны поддержанием благосостояния своего генома. Дело в том, что даже если в генах родителя возникнут вредоносные мутации, есть шанс, что потомкам он передаст хромосому, которая по счастливой случайности будет собрана из «здоровых» частей. Так отбраковывается немалая доля вредоносных генетических изменений. И, наоборот, если по случайности благоприятная мутация возникает среди не самых удачных генетических вариантов, есть шанс, что она отделится от окружения и унаследуется без вредного балласта. В общем, половое размножение — это залог здоровья и благополучия генома.
А как же без этих преимуществ обходятся организмы, которые не размножаются половым путем? Оказывается, если они живут при достаточной плотности (например, в биопленках), их геном по своей мозаичности будет неотличим от генома вида, размножающегося половым путем.
Возможности бактерий по передаче генов намного шире наших. Бактерия может поделиться генами не только со своими потомками, а вообще практически с любой другой бактерией. Передача генов тому, кто не является потомком особи, называется их горизонтальным переносом [1]. Он крайне запутывает филогенетические связи между таксонами и иногда вообще размывает понятие «вид»: у бактерий часто встречаются гены представителей не только другого вида, но иногда даже другого домена (например, горизонтальный обмен ДНК активно идет между термофилами — бактериями и археями) . Поэтому при построении филогенетических деревьев для бактерий и архей ученые испытывают определенные трудности. По-видимому, воссоздать единственное древо для всех прокариот просто нельзя — можно только строить дендрограммы по отдельным генам, которые довольно свободно перемещаются между представителями разных видов и даже царств (рис. 1).
Бактерии настолько «аморальны», что могут обмениваться ДНК вообще много с кем — не ограничиваясь собственным доменом. Горизонтальный генетический перенос между бактериями и археями повинен в формировании крупных ветвей филогенетического древа архей — «Закинули археи эволюционный невод и вытянули...» [2]. Но даже границы прокариотического мира оказались прозрачными для бактериальных генов: паразитическая бактерия вольбахия, например, способна встраивать чуть ли не весь свой генетический скарб в хромосомы насекомых и червей — «Животные обмениваются генами с паразитическими бактериями» [3]. Подобные эксцессы, видимо, изредка случаются и в «пуританском» мире эукариот: в частности, несколько миллионов лет назад предусмотрительная тля позаимствовала гены биосинтеза каротиноидов у грибов. А что, и витаминчик всегда при тебе, и от ос с божьими коровками какая-никакая маскировка: «Гены для синтеза каротиноидов тли получили от грибов» [4]! — Ред.
Проблема эволюционного подхода в микробиологии XX века обсуждается в статье «Эволюция между молотом и наковальней, или как микробиология спасла эволюцию от поглощения молекулярной биологией» [5], а картина мира вообще на заре биологической эволюции — в замечательном эссе «К вопросу о происхождении жизни» [6] — Ред.

Рисунок 1. Дерево жизни. Всё живое согласно классификации К. Вёзе делят на три домена (надцарства) — эукариоты, археи и бактерии. Последние два до 1970-х годов объединяли в единое царство Дробянки (лат. Monera). От редакции: об изменениях, введенных Вёзе, кажется, знают далеко не все авторы российских школьных учебников и «практических подготовок к ЕГЭ» по биологии, учителя и тем более их подопечные. Тип Monera ввел в биологическую систематику Э. Геккель в 1866 году, до царства тип дорос в 1925-м. На дворе год 2015 — ну кончились давно дробянки! Вместе с особым статусом в этом ретроцарстве цианобактерий, по-старинке — сине-зеленых водорослей, которые с водорослями (растениями, эукариотами) филогенетически не имеют ничего общего, поскольку они — обычные бактерии.
рисунок с сайта paramisonenigmas.wordpress.com
Каким бы свободным ни был генетический обмен между бактериями, всё же даже в рамках одного сообщества удается обнаружить группы особей, более похожих друг на друга, чем на остальных. Поскольку людям хочется видеть во всём смысл, они предположили, что каждая группа, представители которой генетически особенно сходны, приспособлена к жизни в определенной микронише. Поэтому такое разделение на группы и сохраняется из поколения в поколение. Действительно, подобные устойчивые «подвиды» удалось обнаружить у бактерий Prochlorococcus [7]. Но это — свободноживущие микроорганизмы, а как обстоит дело в биопленках — ведь, в принципе, в них тоже могут возникать разные микроусловия, способствующие выделению «подвидов»?
Ученые исследовали геномы цианобактерий Synechococcus, образующих биопленки [8], [9]. Анализировался определенный набор локусов каждого генома — на предмет частоты вариаций. Потом исследователи определяли, насколько сходны частоты аллелей в разных сайтах ДНК и как такая корреляция меняется с расстоянием между аллелями. Если предположить, что в бактериальной популяции существуют обособленные «подвиды», то частоты аллелей разных генов внутри подвида должны коррелировать, даже если аллели находятся в молекуле ДНК далеко друг от друга. Однако оказалось, что корреляция резко убывает с расстоянием на ДНК. Подобная закономерность характерна для геномов организмов, размножающихся половым путем: чем дальше там располагаются друг от друга генетические варианты, тем ниже вероятность встретить их в одной и той же хромосоме. Объясняется это просто: чем дальше разнесены на ДНК две мутации, тем больше шансов, что обмен участками хромосом произойдет между ними, и они разойдутся. На разных хромосомах существ, которым свойственно половое размножение, мутации располагаются в самых разных комбинациях, и лишь в очень редких случаях повторяется одна и та же последовательность (набор) мутаций. Такие же следы интенсивной перетасовки генома обнаружились и в биопленках цианобактерий. Получается, что эти микроорганизмы совсем не страдают от отсутствия полового размножения.

В таком мозаичном геноме разные локусы подвергались различному давлению отбора. Авторы оценивали давление отбора исходя из корреляций между редкими и более распространенными вариантами. В отсутствие отбора связаны друг с другом будут только частоты распространенных вариантов — просто по случайности, поскольку их много. А если коррелировать будут и частоты редких аллелей (что вряд ли произойдет случайно) — значит, такие аллели только недавно возникли и еще не успели рекомбинировать. В таком случае можно будет заключить, что редкие аллели — новые, еще только подвергающиеся отбору, а более распространенные — старые и уже отобранные. Для разных исследованных локусов давление отбора оказалось неодинаковым, как и у организмов, пользующихся преимуществами полового размножения. В итоге никаких «подвидов» или экотипов в биопленках не обнаружилось, все составляющие их организмы имели одинаково перетасованные геномы и обменивались генетическим материалом безо всяких ограничений (рис. 2).
Мы, конечно, знали, что горизонтальный перенос помогает прокариотам не вырождаться генетически и отчасти заменяет им половое размножение. Но теперь становится очевидным, что он может вообще не уступать половому размножению по качеству перетасовки генов. Правда, в этом случае для эффективного обмена ДНК, по всей видимости, важна высокая плотность популяции. В результате члены достаточно плотного бактериального сообщества приобретают мозаичные геномы, которые очень пластичны — быстро меняются и не сохраняют выраженных адаптаций к определенным микронишам. Зато сообщество в целом становится, во-первых, очень гибким (подходящие геномы могут быстро собираться из отдельных кусков), а во-вторых, защищенным от накопления вредных изменений (неблагоприятные мутации легче отбраковывать благодаря рекомбинации).
Литература
- Мобильные генетические элементы прокариот: стратификация «общества» бродяжек и домоседов;
- Закинули археи эволюционный невод и вытянули...;
- Элементы: «Животные обмениваются генами с паразитическими бактериями»;
- Элементы: «Гены для синтеза каротиноидов тли получили от грибов»;
- Эволюция между молотом и наковальней, или Как микробиология спасла эволюцию от поглощения молекулярной биологией;
- К вопросу о происхождении жизни;
- Новый вид ингибирования ферментативной активности;
- Michael J. Rosen, Michelle Davison, Devaki Bhaya, Daniel S. Fisher. (2015). Fine-scale diversity and extensive recombination in a quasisexual bacterial population occupying a broad niche. Science. 348, 1019-1023;
- Michael M. Desai, Aleksandra M. Walczak. (2015). Flexible gene pools. Science. 348, 977-978.



Комментарии
0Чтобы оставить комментарий, необходимо
войти