Опасные связи. Новый взгляд на происхождение эукариотических химер, подмявших под себя весь мир
10 декабря 2020
Опасные связи. Новый взгляд на происхождение эукариотических химер, подмявших под себя весь мир
- 4294
- 0
- 6
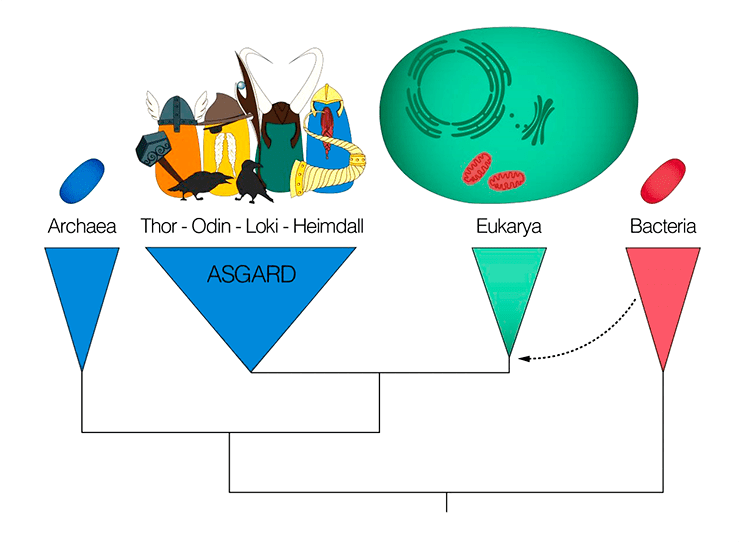
Домен Асгардархеи, в который входят организмы из группы Lokiarchaeota, назван в честь богов скандинавской мифологии и является сестринской группой эукариот
-
Авторы
-
Редакторы
Статья на конкурс «Био/Мол/Текст»: Происхождение эукариот — самое масштабное событие в истории эволюции со времен возникновения жизни. Это огромный скачок во внутренней организации клетки, который позволил живым организмам в дальнейшим перейти к истинной многоклеточности. Но до недавнего времени мы мало что знали про этот переход — каким образом могла возникнуть такая сложная структура, как ядро? Ученые выдвигали на этот счет различные гипотезы, но ни одна из них не объясняла всего процесса. Лишь в последние годы появились более стройные предположения, претендующие на разгадку тайны происхождения эукариот.

Конкурс «Био/Мол/Текст»-2020/2021
 Эта статья участвовала в конкурсе «Био/Мол/Текст»-2020/2021 в номинации «Свободная тема» и будет опубликована в журнале «Наука и жизнь».
Эта статья участвовала в конкурсе «Био/Мол/Текст»-2020/2021 в номинации «Свободная тема» и будет опубликована в журнале «Наука и жизнь».
Генеральный партнер конкурса — ежегодная биотехнологическая конференция BiotechClub, организованная международной инновационной биотехнологической компанией BIOCAD.

Спонсор конкурса — компания SkyGen: передовой дистрибьютор продукции для life science на российском рынке.
Спонсор конкурса — компания «Диаэм»: крупнейший поставщик оборудования, реагентов и расходных материалов для биологических исследований и производств.
«Книжный» спонсор конкурса — «Альпина нон-фикшн»
Суть проблемы
Происхождение эукариот остается одной из самых больших загадок в современной биологии. Прокариоты (организмы без ядра) и эукариоты (организмы с ядром) различаются между собой организацией генома — у эукариот ДНК заключена в специализированном компартменте клетки, обособленном от всего остального пространства мембраной, тогда как у прокариот она находится прямо в цитоплазме. Это небольшое отличие сыграло важную эволюционную роль. Более совершенная регуляция генома позволила эукариотам защититься от спонтанных мутаций, которые неизбежно возникают, когда генетическая информация хранится в области активного метаболизма — цитоплазме [1], [2].
Появление эукариотической клетки с ее ядром, эндомембранной системой и мембранными органеллами представляет собой качественный сдвиг во внутреннем устройстве, по сложности превосходящий всё, что можно увидеть у прокариот. Но что именно позволило эукариотам приобрести ядро, сделав переход на совершенно иной уровень клеточной организации?
Появление ядра неразрывно связано с другим процессом в эволюции эукариот — симбиозом. Предок эукариот вступил в симбиотические отношения с альфа-протеобактерией, будущей энергетической станцией клетки — митохондрией. Но несмотря на то, что самой идее возникновения митохондрий уже десятки лет, каждый год ученые обнаруживают нечто, что заставляет по-новому взглянуть на этот союз и на происхождение эукариот в целом [3].
Первые шаги к пониманию

Рисунок 1. Линн Маргулис, создательница современной версии теории симбиогенеза
В далеком 1967 году в научном журнале Journal of Theoretical Biology («Журнал теоретической биологии») вышла статья On the origin of mitosing cells («О происхождении клеток, делящихся митозом»). Автором этой статьи была Линн Маргулис, женщина, которая своими исследованиями произвела настоящую сенсацию в научном сообществе (рис. 1).
Еще со времен Чарльза Дарвина ученые считали, что единственным путем эволюции является дивергенция, то есть расхождение видов. В то время не вызывало никаких сомнений, что один вид живых существ с течением времени может лишь разделяться на множество других. Однако работы Линн Маргулис показали, что механизм некоторых эволюционных событий был принципиально иным: оказалось, что эукариоты образовались путем слияния разных эволюционных ветвей, а значит, эволюция сводится не к дивергенции — иногда разделившиеся довольно давно виды могут объединяться снова. Как говорил на заре своей политической карьеры и совершенно по другому вопросу Владимир Ленин: «Для того чтобы объединиться, необходимо сначала решительно и определенно размежеваться».
Основная идея эндосимбиотической гипотезы гласит, что митохондрии произошли от бактерий, и сейчас это уже не подвергается сомнению. Однако долгое время — до обнаружения ДНК в митохондриях — ее считали в лучшем случае сомнительной. Борьба была долгой и упорной, но вот, когда гипотеза была принята, возникли новые вопросы. Кем же был этот загадочный предок эукариот, вступивший в симбиоз с митохондриями? И что подтолкнуло эти организмы к симбиозу?
Новый источник энергии
Ответить на последний вопрос достаточно просто — появление эукариот совпадает по времени с крупнейшей за всю историю Земли геофизической перестройкой.
В древнейшие времена доминирующей формой жизни на нашей планете были цианобактерии. Именно они первыми научились кислородному фотосинтезу, и, получая энергию из неорганических веществ и солнечного света, цианобактерии выделяли в атмосферу ненужный побочный продукт фотосинтеза — кислород. Примерно 2,45 млрд лет назад содержание кислорода в земной атмосфере достигло опасно высокого уровня. Для всех живущих в то время организмов — которые, разумеется, были анаэробами, — кислород являлся сильнейшим ядом, что связано с образованием активных форм кислорода (АФК), повреждающих биомолекулы [4]. Аэробы отличаются наличием защитной антиоксидантной системы, способной к обезвреживанию АФК.
В итоге из-за слишком высокого содержания кислорода в атмосфере началось первое в истории массовое вымирание [5], [6]. В это неспокойное время в выигрыше остались альфа-протеобактерии, умевшие эффективно использовать кислород для получения энергии. Помимо них выжил и загадочный предок эукариот, который пошел по более легкому, но, вместе с тем, более изощренному пути, вступив в симбиоз с альфа-протеобактерией. В результате этого союза образовалась «химерная» клетка, получившая возможность дышать кислородом и породившая новую ветвь эволюции, из которой возникли эукариоты. Похожая ситуация произошла с цианобактериями — из них образовались хлоропласты, дав некоторым эукариотам возможность фотосинтезировать (и породив этим ветвь растений) [7], [8].
Сами по себе митохондрии и хлоропласты делятся независимо от клетки, хранят свою собственную генетическую информацию и получают от клетки большое количество необходимых веществ, но, переложив столько функций на клетку-хозяина, они теряют автономию и уже не могут жить отдельно от нее. Такой союз называется синтрофией — типом симбиотического сосуществования, в котором один вид живет за счет продуктов метаболизма другого вида.
Гипотеза фагоцитоза
Переход от совместного сосуществования к эндосимбиозу — весьма серьезный шаг для клетки, который предполагает большие структурные изменения. Чтобы объяснить происхождение митохондрий была выдвинута гипотеза фагоцитоза. В своем классическом варианте она гласит: предки современных эукариот, значительно отличавшиеся и от бактерий, и от архей, самостоятельно приобрели большинство признаков, свойственных эукариотам — цитоскелет, систему внутренних мембран, и, наконец, ядро. Позже они захватили альфа-протеобактерию, то есть, будущую митохондрию.
Кто приручил митохондрию?
Однако сейчас ореол загадочности, окружавший нашего предка и мешавший разглядеть то, что лежало под самым носом, развеялся. Первый общий предок эукариот (FECA) — самый древний организм, от которого произошли все эукариоты, — являлся, судя по всему, самой обычной археей. Эта идея не сразу была принята научным сообществом — потребовалось немало времени, чтобы ее хотя бы начали рассматривать всерьез [9], [10].
Но прежде чем подробнее изучить предка эукариот, давайте посмотрим на временную эволюционную линию (рис. 2). LUCA (last universal common ancestor) — это последний общий предок всех живых организмов. Ископаемых остатков LUCA, конечно, не сохранилось, поэтому его можно изучать только путем сравнения геномов, и, судя по этим данным, LUCA впоследствии разделился на два домена — бактерий и архей. Приобретя митохондрию и ядро, одна из архей — FECA — превратилась в LECA (last eukaryotic common ancestor) — полноценного эукариота. Эндосимбиоз должен был предшествовать LECA, поскольку сейчас не существует эукариот, полностью лишенных митохондрий (некоторые утратили митохондрии вторично). Следует отметить, что LECA — не первый полноценный эукариот, а последний общий предок всех современных эукариотов.

Рисунок 2. Поворотные точки в эволюции эукариот. LUCA — последний общий предок живых организмов; FECA — первый общий предок эукариот; LECA — последний общий предок эукариот.
Когда исследователи принялись изучать геном архей группы Lokiarchaeota, неожиданно оказалось, что эти морские донные микроорганизмы — наши давно потерянные родственники: вместе с эукариотами они образуют монофилетическую кладу, то есть группу организмов с общим предком [11]. Эти данные были получены методом молекулярной филогенетики .
Молекулярная филогенетика — способ установления родственных связей между организмами на основании изучения структуры ДНК, РНК и белков. Для филогенетических исследований часто используют ген 16S рРНК — последовательность этой молекулы содержит консервативные (16S рРНК из эволюционно далеких видов бактерий имеют сходные участки последовательности и функции) и вариабельные (разнящиеся от вида к виду) участки.
В 2019 году привередливую культуру локиархеот впервые удалось вырастить в лабораторных условиях. Это был сложный и трудоемкий процесс, занявший у исследователей целых 12 лет, потому что, как выяснилось, эти археи не могут расти в виде монокультуры (то есть в культуре, состоящей из одного вида). Для создания оптимальной среды пришлось соорудить биореактор, который поддерживал температуру 10 °С, а также низкий уровень кислорода и питательных веществ. Через пять лет работы реактора в нем вырос пестрый конгломерат из бактерий и архей, причем археи доминировали и среди них оказался весьма примечательный микроорганизм — P. syntrophicum. Еще семь лет понадобилось на то, чтобы нарастить культуру в достаточном количестве — археи этой группы размножаются чрезвычайно медленно, удвоение клетки занимает от 14 до 25 дней [12], [13]. Наконец, количество микроорганизмов в биореакторе достигло пригодных для изучения значений. И вот у нас появилась возможность воочию увидеть пусть не своего прямого предка, но достаточно близкий к нему организм, и выращенная японскими учеными (с поистине азиатским усердием) архея нас не разочаровала.
Детальное исследование локиархеот показало, что органоидов они лишены, но от них могут отпочковываться мембранные везикулы, а кроме того, эти археи формируют особые мембранные выросты — протрузии (рис. 3). Они позволяют локиархеотам расти в тесном контакте с археями рода Methanogenium, которые потребляют вещества, препятствующие росту локиархеот [8], то есть находятся в тесных синтрофических отношениях. Термин синтрофия уже встречался нам, когда речь шла о митохондриях.

Рисунок 3. Протрузии P. syntrophicum. Протрузии — мембранные выросты архей, которые позволяют им жить в синтрофных отношениях с другими видами архей.
Но почему то, что археи, близкие к нашим родственникам, смогли образовать какие-то выросты — важно? Дело в том, что этот небольшой факт позволяет заполнить сразу несколько белых пятен, которые до сих пор так резали глаза при взгляде на эволюционную историю эукариот. Во-первых, отсутствие фагоцитоза и наличие протрузий дает более реалистичную альтернативу гипотезе фагоцитоза, которая, пусть и является общепринятой в научном сообществе, не лишена своих недостатков. Во-вторых, это, наконец, проливает свет на способ образования ядра. Но обо всем по порядку.
Никто никого не ел?
Из всех живых организмов лишь эукариоты обладают фагоцитозом, но не путаем ли мы причину со следствием, утверждая, что он был причиной появления эукариот в таком виде, в каком мы их знаем сейчас? Гипотеза фагоцитоза гласит: FECA — ранний предок эукариот — поглотил бактерию, но по каким-то причинам не переварил ее, а стал использовать для получения энергии. Доказать это экспериментально так и не удалось, однако до недавнего времени она давала ответы на большинство вопросов.
Но всё же не на все. И вот культивирование локиархеот показало нам альтернативный способ совместного существования — при помощи протрузий. С тех пор гипотеза синтрофии получала все больше и больше подтверждений.
В первую очередь в ее пользу говорит то, что до появления митохондрий фагоцитоз был энергетически невыгоден клетке. Этот процесс очень энергозатратен — добычу требуется догнать, поймать и переварить. Без митохондрий на этот процесс энергии тратится больше, чем клетка способна в принципе получить в результате поглощения пищи, полученной таким способом [14]. А ведь средняя эукариотическая клетка потребляет примерно в 5000 раз больше энергии, чем прокариотическая [15], [16].
Палеонтология, сколь бы мало она не могла сказать нам о жизни микроорганизмов, тоже ставит под сомнение раннее появление фагоцитоза. Надежные свидетельства его существования появляются в палеонтологической летописи около 1 млрд лет назад. Между тем, LECA, ближайший общий предок всех современных эукариот (то есть организм, от которого отделились все современные эукариоты) жил примерно 1,6–1,8 млрд лет назад — то есть был уже вполне сформированным эукариотом, не обладавшим фагоцитозом [17], [18].
Все это дает основания рассматривать синтрофную гипотезу появления эукариот наравне с наиболее принятой сейчас — гипотезой фагоцитоза [19]. Более того, она предлагает нам возможный ответ на один из наиболее важных вопросов в эволюционной истории жизни.
Загадка происхождения ядра. Вывернуться наизнанку, чтобы выжить
Несмотря на огромный прогресс цитологии и молекулярной биологии, в истории происхождения эукариот, как мы выяснили, до сих пор хватает пробелов. Мало того, мы до сих пор не знаем, как возникла самая главная часть эукариотической клетки — ядро!
Сегодня существуют несколько гипотез, которые попытались объяснить происхождение ядра.
Первая гипотеза называется синтропной моделью и предполагает, что ядро появилось в результате симбиоза археи и бактерии. Согласно ей, древняя архея проникла в бактерию, где впоследствии редуцировалась до клеточного ядра эукариот [20].
Вторая гипотеза говорит о том, что бактерия эволюционировала в эукариота без эндосимбиоза и опирается лишь на существование бактерий рода Planctomycetes, имеющих структуры, напоминающие ядро [21].
Третья гипотеза — это гипотеза вирусного эукариогенеза, которая предполагает, что ядро возникло вследствие заражения прокариотической клетки вирусом. По одной версии, ядро возникло при поглощении клеткой большого ДНК-содержащего вируса [22], по другой — эукариоты произошли от древних архей, уже инфицированных поксвирусами [23].
Четвертая гипотеза, названная экзомембранной, утверждает, что ядро произошло от одиночной клетки, выработавшей вторую внешнюю мембрану. Первичная мембрана превратилась в ядерную и в ней появились поровые структуры для транспорта синтезированных внутри компонентов. Однако большой поддержкой она тоже не пользуется, поскольку предполагает независимое происхождение прокариот и эукариот [24].
Ни одна из этих гипотез не является общепризнанной, каждая имеет достаточно серьезные противоречия. Однако не все так безнадежно, как может показаться. В 2014 году вышла статья, в которой исследователи выдвинули новую гипотезу происхождения ядра — гипотезу, получившую название inside-out, то есть «снаружи—внутрь», или «наизнанку» (рис. 4). Во многом своим происхождением она обязана развитию экзомембранной гипотезы, но имеет от нее ряд отличий. Предположение о происхождении клетки «наизнанку» примечательно тем, что не опирается на наличие фагоцитоза у FECA (которого у него, судя по всему, и не было), что позволяет разрешить часть существовавших ранее трудностей. Согласно этой гипотезе, ядро произошло от одной клетки, которая в процессе эволюции образовала вторую внешнюю клеточную мембрану, а прежняя после этого стала ядерной [25].

Рисунок 4. Последовательные этапы эволюции первого общего предка эукариот (FECA) согласно гипотезе inside-out. а — Архейный предок с единственной мембраной и богатой гликопротеинами клеточной стенкой (S-слой) взаимодействует с альфа-протеобактериями — будущими митохондриями. б — Клетка образует выступы — протрузии, — которые облегчают обмен метаболитами с альфа-протеобактериями. в — Увеличение площади контакта между симбионтами приводит к росту протрузий и возможной потере S-слоя. г — Протрузии в дальнейшем развиваются в ядерный комплекс, который, благодаря постепенной потере S-слоя, физически контактирует с мембраной клетки. д — С расширением протрузий, которые охватывают симбионтов, они постепенно перемещаются в цитоплазму. е — Наконец, слияние протрузий соединяет цитоплазматические компартменты воедино. Такой переход изолирует эндоплазматический ретикулум от внешней среды, что одновременно помогает развитию везикулярного транспорта и устанавливает вертикальную передачу митохондрий, а это приближает нашего гипотетического предка к клетке с современной эукариотической организацией.
Как мы уже знаем, археи способны создавать клеточные выпячивания — протрузии, но при этом не обладают фагоцитозом. Именно на этом и основывается гипотеза inside-out. Ее авторы предполагают, что эукариоты произошли от клетки, которая расширила свои протрузии, а они, сливаясь, дали начало цитоплазме и системе внутренних мембран. Согласно гипотезе inside-out, внешняя ядерная мембрана, плазматическая мембрана и цитоплазма произошли из внеклеточных выступов, тогда как эндоплазматический ретикулум представляет собой промежутки между пузырьками. Митохондрии первоначально были захвачены в эндоплазматический ретикулум, но позже проникли через его мембрану, попав в цитоплазму. Согласно этой модели заключительным этапом эукариогенеза было формирование непрерывной плазматической мембраны, которая закрывала эндоплазматический ретикулум снаружи.
Подобные примеры сосуществования архей и бактерий известны и в настоящее время — например, группа таумархиот, образующая эктосимбиоз с гамма-протеобактериями [29].

Рисунок 5. Микрофотография двух клеток археи Candidatus Giganthauma karukerense, окруженных эктосимбиотическими γ-протеобактериями
Заключение
Что же в итоге? Синтрофная гипотеза гласит о том, что предок эукариот был менее прожорливым, чем мы привыкли считать. Вместо поедания бактерий он как бы «обнимал» их своими протрузиями, и сеть выростов в дальнейшем расширялась, создавая ячейки для бактерий-симбионтов и отделяя оболочку будущего ядра. Так постепенно, шаг за шагом и формировалась эукариотическая клетка. Эта гипотеза представляет собой свежую альтернативу гипотезе фагоцитоза, предполагающей, что предок эукариот поглотил и внедрил в себя альфа-протеобактерию.
Разумеется, в научном мире тяжело менять устоявшиеся концепции, особенно когда они укоренились настолько глубоко. Гипотеза фагоцитоза известна давно и принята повсеместно, поэтому изменить привычный взгляд на происхождение митохондрий непросто, но в свете последних открытий ее явно нужно пересмотреть. Гипотеза синтрофии позволяет разрешить ряд давних проблем, с которыми не справилась гипотеза фагоцитоза: она согласуется с имеющимися данными о наших предках и отлично стыкуется с гипотезой происхождения ядра inside-out, не имея при этом противоречий, связанных с палеонтологией или энергетикой клетки.
Но не стоит забывать и о том, что дьявол кроется в деталях. Мы до сих пор можем лишь предполагать, какими веществами обменивались в синтрофическом союзе FECA и альфа-протеобактерия и даже еще не выяснили, чем обмениваются локиархеи со своими симбионтами. Многое только предстоит выяснить, но если гипотеза и окажется неверной, наверняка во время ее проверки удастся совершить массу научных открытий.
Литература
- Charles F. Baer, Michael M. Miyamoto, Dee R. Denver. (2007). Mutation rate variation in multicellular eukaryotes: causes and consequences. Nat Rev Genet. 8, 619-631;
- Кассимерис Л., Окс Р., Льюин Б. Клетки по Льюину. «Лаборатория знаний», 2016. — 1056 с.;
- Anthony M. Poole, David Penny. (2007). Evaluating hypotheses for the origin of eukaryotes. Bioessays. 29, 74-84;
- Федоров А.А. Жизнь растений (том 1). 2012;
- Margulis L. and Sagan D. Microcosmos: four billion years of microbial evolution. University of California Press, 1997. — 300 p.;
- Timothy W. Lyons, Christopher T. Reinhard, Noah J. Planavsky. (2014). The rise of oxygen in Earth’s early ocean and atmosphere. Nature. 506, 307-315;
- Patrick J. Keeling. (2013). The Number, Speed, and Impact of Plastid Endosymbioses in Eukaryotic Evolution. Annu. Rev. Plant Biol.. 64, 583-607;
- Nick Lane. (2017). Serial endosymbiosis or singular event at the origin of eukaryotes?. Journal of Theoretical Biology. 434, 58-67;
- Purificación López-García, David Moreira. (2015). Open Questions on the Origin of Eukaryotes. Trends in Ecology & Evolution. 30, 697-708;
- C. J. Cox, P. G. Foster, R. P. Hirt, S. R. Harris, T. M. Embley. (2008). The archaebacterial origin of eukaryotes. Proceedings of the National Academy of Sciences. 105, 20356-20361;
- Anja Spang, Jimmy H. Saw, Steffen L. Jørgensen, Katarzyna Zaremba-Niedzwiedzka, Joran Martijn, et. al.. (2015). Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature. 521, 173-179;
- Hiroyuki Imachi, Masaru K. Nobu, Nozomi Nakahara, Yuki Morono, Miyuki Ogawara, et. al.. (2020). Isolation of an archaeon at the prokaryote–eukaryote interface. Nature. 577, 519-525;
- Собственной персоной: ученым удалось вырастить в лаборатории вероятного предка эукариот;
- William F. Martin, Aloysius G. M. Tielens, Marek Mentel, Sriram G. Garg, Sven B. Gould. (2017). The Physiology of Phagocytosis in the Context of Mitochondrial Origin. Microbiol Mol Biol Rev. 81;
- Nick Lane. (2011). Energetics and genetics across the prokaryote-eukaryote divide. Biology Direct. 6, 35;
- От сложного к простому: трудности симбиогенеза;
- L. W. Parfrey, D. J. G. Lahr, A. H. Knoll, L. A. Katz. (2011). Estimating the timing of early eukaryotic diversification with multigene molecular clocks. Proceedings of the National Academy of Sciences. 108, 13624-13629;
- Элементы: «Когда появился фагоцитоз?»;
- Элементы: «Предложена новая гипотеза происхождения эукариот»;
- Purificación López-García, David Moreira. (2006). Selective forces for the origin of the eukaryotic nucleus. Bioessays. 28, 525-533;
- John A. Fuerst. (2005). INTRACELLULAR COMPARTMENTATION IN PLANCTOMYCETES. Annu. Rev. Microbiol.. 59, 299-328;
- Philip John Livingstone Bell. (2001). Viral Eukaryogenesis: Was the Ancestor of the Nucleus a Complex DNA Virus?. Journal of Molecular Evolution. 53, 251-256;
- Masaharu Takemura. (2001). Poxviruses and the Origin of the Eukaryotic Nucleus. J Mol Evol. 52, 419-425;
- Albert D. G. de Roos. (2006). The Origin of the Eukaryotic Cell Based on Conservation of Existing Interfaces. Artificial Life. 12, 513-523;
- David A Baum, Buzz Baum. (2014). An inside-out origin for the eukaryotic cell. BMC Biol. 12;
- A. M. Poole, S. Gribaldo. (2014). Eukaryotic Origins: How and When Was the Mitochondrion Acquired?. Cold Spring Harbor Perspectives in Biology. 6, a015990-a015990;
- Natalya Yutin, Maxim Y Wolf, Yuri I Wolf, Eugene V Koonin. (2009). The origins of phagocytosis and eukaryogenesis. Biology Direct. 4, 9;
- Joran Martijn, Thijs J.G. Ettema. (2013). From archaeon to eukaryote: the evolutionary dark ages of the eukaryotic cell. Biochemical Society Transactions. 41, 451-457;
- Félix Muller, Terry Brissac, Nadine Le Bris, Horst Felbeck, Olivier Gros. (2010). First description of giant Archaea (Thaumarchaeota) associated with putative bacterial ectosymbionts in a sulfidic marine habitat. Environmental Microbiology. 12, 2371-2383;
- Eelco C. Tromer, Jolien J. E. van Hooff, Geert J. P. L. Kops, Berend Snel. (2019). Mosaic origin of the eukaryotic kinetochore. Proc Natl Acad Sci USA. 116, 12873-12882.



Комментарии
0Чтобы оставить комментарий, необходимо
войти