Атлас гистоновых модификаций в раннем митозе
26 мая 2026
Атлас гистоновых модификаций в раннем митозе
- 546
- 0
- 3
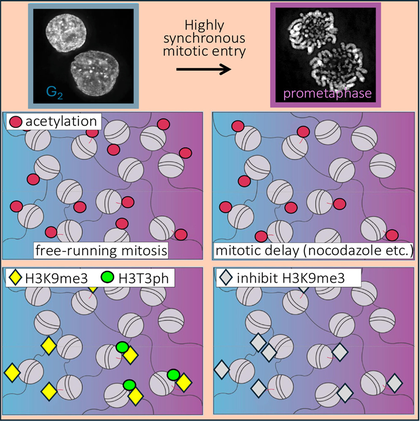
Гистоновые модификации были измерены во время очень синхронного раннего митоза (сверху). В этой системе гистоны не деацетилируются при входе клеток в митоз, в отличие от традиционного блока клеток в митозе нокодазолом (в центре). Гистоновая метка H3T3ph (фосфорилирование гистона H3 по треонину 3) является гетерохроматиновой, и ее навешивание зависит от главной гетерохроматиновой метки H3K9me3 (снизу).
Рисунок в полном размере.
графическая аннотация к статье A time-resolved atlas of histone modifications during mitotic entry [10]
-
Автор
-
Редакторы
Темы
В митозе хромосомы конденсируются в белковые остовы с исходящими из них петлями хроматина: объем хромосом уменьшается в 2–3 раза, а гистоны фосфорилируются и, в определенных системах, деацетилируются. Однако до недавнего времени отсутствовала информация о кинетике навешивания гистоновых модификаций в раннем митозе. Более того, на базе информации при блоке клеток в митозе нокодазолом возникла гипотеза, что деацетилирование гистонов в митозе ведет к компактизации хромосом. Однако полного подтверждения или опровержения этой гипотезы до недавнего времени не было. Эти пробелы устранила международная команда из Мюнхена и Эдинбурга, которая составила атлас гистоновых модификаций в раннем митозе, а также показала, что в плане деацетилирования гистонов система с нокодазолом ведет к артефактам.
При делении клетки в ней происходит множество внутренних перестроек. Хромосомы конденсируются и приобретают палочковидную форму, а их объем уменьшается в 2–3 раза по сравнению с интерфазным [1] . Ядерная оболочка разрушается, и формируется веретено деления. Хромосомы выстраиваются в метафазную пластинку у экватора, а затем микротрубочки веретена деления растаскивают их в анафазе к полюсам клетки, которая в итоге делится на две.
Больше про митоз и клеточный цикл можно прочитать в статье спецпроекта «Эпигенетика»: «Процессы и Эпигеном» [2].
При изменении облика хромосом с ними, в свою очередь, тоже происходят перестройки . Топологически ассоциированные домены — контактирующие участки на картах Hi-C, представляющие собой небольшие глобулы хроматина порядка мегабазы, — исчезают, а каждая длинная и запутанная пара молекул ДНК с нуклеосомами перевыпетливается конденсином, причем петли исходят из белкового спирального остова, образованного этим самым конденсином и топоизомеразой II [3]. Конденсин I генерирует петли внутри петель конденсина II. У митотических хромосом образуется оболочка из РНК и белков, самый известный из которых — раковый маркер Ki-67.
Больше про интерфазную укладку генома в пространстве можно узнать в статье «Биомолекулы»: «Новый взгляд на геном: не просто цепочка генов, а трехмерная сеть, интегрирующая функциональные домены ядра» [1].
Помимо образования определенной архитектуры, происходит уменьшение объема хромосом, и похоже, этот процесс не зависит от конденсации хромосом конденсином. Если убрать конденсин прямо перед входом клеток в митоз, то хромосомы превращаются в одну сплошную массу, теряя палочковидную структуру. Однако объем этой массы равен сумме объемов отдельных палочек в контрольных клетках [4]. После этого открытия, а также in vitro экспериментов, в которых искусственный хроматин из митотических гистонов агрегировал сильнее, чем из интерфазных [5], несколько лет думали, что уменьшение объема хроматина в митозе происходит за счет изменения гистоновых модификаций, конкретно — митотического деацетилирования гистонов . Эту гипотезу, а также детали изменений митотического фосфорилирования гистонов, проверяли ученые под руководством Акселя Имхофа и Билла Эрншоу в коллаборации между Мюнхенским и Эдинбургским университетами.
Больше про гистоновые модификации можно узнать в в статьях спецпроекта «Эпигенетика»: «Молекулы и эпигеном» [6] и Ядро и эпигеном» [7].
То, что гистоны в митозе фосфорилируются, известно давно. Киназа Аврора Б модифицирует фосфатными остатками H3S10 (серин 10 гистона H3) и H3S28 (серин 28 гистона H3). Киназа Haspin модифицирует фосфатами H3T3 (треонин 3 гистона H3). Киназа Bub1 фосфорилирует H2AT120 (треонин 120 гистона H2A), и вместе с H3T3ph эта метка важна для регуляции митоза определенными белковыми комплексами. Однако все это было показано либо иммунофлуоресценцией, которую трудно сделать количественной, либо масс-спектрометрией [8] при накоплении клеток в прометафазе нокодазолом, то есть в системе без временного разрешения.
В распоряжении лаборатории Билла Эрншоу уже несколько лет была клеточная линия куриной лимфомы DT40, чья киназа CDK1, запускающая митоз, имела точечную мутацию в активном центре [3], [9]. Благодаря этой мутации ее можно было заблокировать в неактивном состоянии аналогом АТФ 1NM-PP1. При этом клетки накапливались в фазе G2, а при быстрой смене среды (удалении ингибитора) они входили в митоз настолько синхронно, что более 90% клеточной популяции находилось на одной и той же стадии профазы. Время, за которое они добирались до стадии прометафазы, составляло минуты.
Действительно, давайте рассмотрим эту систему в действии в обсуждаемой статье (рис. 1) [10]. В 0,5 минут или 2,5 минуты после входа в митоз клетки находятся в ранней профазе, а ядерная оболочка сохраняется. В 5 или 7,5 минут хроматин уже более конденсирован в поздней профазе, хотя ядерная оболочка все еще не разрушена. В 15 минут хромосомы уже полностью конденсированы, а от ядерной оболочки остались куски в цитоплазме и на хроматине. В 30 минут то, что было ядерной оболочкой, уже диффундировало по всей цитоплазме делящейся клетки.

Рисунок 1. Микрофотографии клеток, вошедших в митоз в течение определенного времени. Ядерная оболочка отмечена иммунофлуоресценцией Ламина Б1 (Lamin B1). Этот ламин также присутствует в небольших количествах внутри ядра.
С такой системой удалось проследить не только общие тенденции изменения гистоновых модификаций, но и провести количественный анализ отдельных меток в каждой точке времени — все благодаря отделу масс-спектрометрии из Мюнхена. Из модификаций фосфорилирования были проанализированы H3S10ph, H3T3ph и H3.1S28ph — у гистона H3 два основных варианта, кодируемых разными генами, и последнюю модификацию удалось измерить только на варианте H3.1. Каждую метку измеряли как процент данной модификации от интенсивности данного пептида со всеми возможными модификациями.
Эти три метки вели себя по-разному. H3S10ph начинало подниматься в 2,5 минуты, и достигало практически максимального значения около 80% в 5 минут, оставаясь высоким на протяжении большей части раннего митоза. H3T3ph значительно поднималось только к пяти минутам, достигало максимального значения около 30% в 7,5–15 минут и потом немного снижалось. H3.1S28ph демонстрировало третий временной паттерн — эта метка значительно поднималась только к 7,5 минутам и достигала пика где-то в 30% к 15–30 минутам. Получается, в митозе есть как минимум три волны фосфорилирования, которые накатывают на хроматин одна за другой с разницей в 2,5 минуты!

Рисунок 2. Доля пептидов с H3S10ph, H3T3ph и H3.1S28ph в каждой измеренной точке времени раннего митоза.

Рисунок 3. Имуннофлуоресценция меток H3T3ph и H3K9me3 в клетках в фазе G2 и в 30 минут раннего митоза. При воздействии хаетоцином (cheatocin) — ингибитором метилтрансферазы, навешивающей H3K9me3, — интенсивность H3T3ph сильно падает. Следовательно, H3K9me3 важно для установления метки H3T3ph.
Рядом с описанными фосфорилирующимися остатками в гистонах находятся лизины, на которые навешиваются метильные метки. В частности, за H3T3 следует H3K4. H3K4me1 (монометилирование) — известная метка энхансеров, а H3K4me2 и me3 — промоторов [11]. Причем, согласно измерениям масс-спектрометрией, последние две метки каждая составляют менее одного процента всего пептида, содержащего H3K4. С одной стороны, это неожиданно — ведь эти метки присутствуют на каждом промоторе! С другой стороны, если подумать, промотор — это очень маленькая область гена, а всего гены занимают только 2% длины ДНК, так что все логично. Три вышеописанные метильные метки, а также H3T3ph, исследователи решили изучить иммунопреципитацией хроматина. Этим методом можно посмотреть, в каких местах линейной ДНК находится, например, данная гистоновая модификация, а также независимо от масс-спектрометрии измерить количество меток на ДНК.
Метки H3K4me1, me2 и me3 оставались стабильными в раннем митозе — они не менялись. Согласуясь с результатами масс-спектрометрии, H3T3ph повышалось. Однако самая интересная вещь выяснилась в распределении метки H3T3ph — она коррелировала с меткой неактивного гетерохроматина и молчащих генов H3K9me3. Ученые решили воздействовать на клетки ингибитором метилтрансферазы, которая навешивает H3K9me3, прямо перед митозом, и H3T3ph почти совсем исчезло (рис. 3)! Получается, H3T3ph не только гетерохроматин-специфичный маркер митоза, но навешивание этой метки направляется главной гистоновой модификацией гетерохроматина.
Больше про гетерохроматин и другие домены ядра можно прочитать в статье спецпроекта «Эпигенетика» «Ядро и эпигеном» [7].
А что же с деацетилированием гистонов — меткой, которая, по идее, должна «рулить» компактизацией митотических хромосом? Исследователи сравнили деацетилирование гистонов в системе с синхронным ранним митозом и системе, которую повсеместно использовали до них — блок клеток в прометафазе нокодазолом. При нокодазоле, как и было показано до этого, гистоны отлично деацетилировались. А вот в системе с синхронным ранним митозом этого практически не происходило (рис. 4)!

Рисунок 4. Изменение ацетилирования в системе с синхронным ранним митозом и при добавлении нокодазола, который блокирует клетки в прометафазе и не дает им прогрессировать в митозе дальше (последний стоблик). Синий цвет обозначает уменьшение, а красный — увеличение. Как видно из тепловой карты, гистоны практически не деацетилируются к 30 минутам раннего митоза по сравнению с тем, что происходит в нокодазоле — там деацетилирование прошло полным ходом!
Начали разбираться, что происходит, попробовали другой блокатор клеток в прометафазе — MG132. Гистоны также деацетилировались, причем пропорционально времени, проведенному клетками в митозе. Этому есть только одно объяснение — в синхронной системе ученых вход в митоз очень быстрый, за минуты. Традиционные методы накопления клеток в митозе, напротив, предполагают длительную инкубацию клеток с блокатором. При этом в большинстве клеток популяции митоз оказывается сильно продлен, что и ведет к деацетилированию гистонов. Получается, гистоны практически не деацетилируются в нормальном митозе! Но что же тогда двигает компактизацию митотических хромосом?
Ученые склоняются к гипотезе, которую изначально опубликовал Кацухиро Маешима [12]. Согласно его экспериментам, в митозе поднимается концентрация ионов магния, не в последнюю очередь за счет повального использования АТФ митотическими киназами и конденсинами — АТФ и Mg2+ находятся в комплексе, и в раннем митозе Mg2+ высвобождается из этого комплекса. Ионы магния преципитируют хроматин, нейтрализуя негативный заряд ДНК и делая хроматин компактнее.
Таким образом, в данной работе [10] удалось не только узнать большее о биологии метки H3T3ph, но и опровергнуть гипотезу деацетилирования гистонов в митозе как драйвера компактизации хромосом, которая была изначально высказана в лаборатории Билла Эрншоу [5] и обсуждалась научным сообществом более пяти последних лет (также см. [13]).
Литература
- Новый взгляд на геном: не просто цепочка генов, а трехмерная сеть, интегрирующая функциональные домены ядра;
- Процессы и эпигеном;
- Johan H. Gibcus, Kumiko Samejima, Anton Goloborodko, Itaru Samejima, Natalia Naumova, et. al.. (2018). A pathway for mitotic chromosome formation. Science. 359, eaao6135;
- Kumiko Samejima, Daniel G. Booth, Hiromi Ogawa, James R. Paulson, Linfeng Xie, et. al.. (2018). Rapid degradation of condensins and 3D-EM reveal chromatin volume is uncoupled from chromosome architecture in mitosis. Journal of Cell Science;
- Alisa Zhiteneva, Juan Jose Bonfiglio, Alexandr Makarov, Thomas Colby, Paola Vagnarelli, et. al.. (2017). Mitotic post-translational modifications of histones promote chromatin compaction in vitro. Open Biol.. 7, 170076;
- Молекулы и эпигеном ;
- Ядро и эпигеном;
- 12 методов в картинках: протеомика;
- Helfrid Hochegger, Donniphat Dejsuphong, Eiichiro Sonoda, Alihossein Saberi, Eeson Rajendra, et. al.. (2007). An essential role for Cdk1 in S phase control is revealed via chemical genetics in vertebrate cells. Journal of Cell Biology. 178, 257-268;
- Natalia Y. Kochanova, Moonmoon Deb, Marco Borsò, Shaun Webb, Ipek Ustun, et. al.. (2026). A time-resolved atlas of histone modifications during mitotic entry. Molecular Cell. 86, 1653-1672.e9;
- The ENCODE Project Consortium. (2012). An integrated encyclopedia of DNA elements in the human genome. Nature. 489, 57-74;
- Kazuhiro Maeshima, Kayo Hibino, Damien F. Hudson. (2018). Condensins under the microscope. Journal of Cell Biology. 217, 2229-2231;
- Matthias Schneider, Benjamin Ertl, Christopher J. Diekmann, Farahnaz Khosrawi, Andreas Weber, et. al.. (2022). Design and description of the MUSICA IASI full retrieval product. Earth Syst. Sci. Data. 14, 709-742.



Комментарии
0Чтобы оставить комментарий, необходимо
войти