Особенности механизмов зрения птиц
18 января 2022
Особенности механизмов зрения птиц
- 6542
- 0
- 9
Глаз птицы по праву считается самым малоизученным. За время эволюции он сохранил особенности зрения наших далеких общих предков, обладает двойными палочками и встроенной системой навигации. Чаще всего под рукой исследователей оказываются глаза цыплят. Но даже глаз обычного цыпленка позволяет приоткрыть секреты восприятия мира наших пернатых собратьев!
Рисунок в полном размере.
-
Автор
-
Редакторы
Статья на конкурс «Био/Мол/Текст»: Птицы — удивительные создания, господствующие в воздухе среди всех хордовых. Численность видов птиц на три тысячи превосходит млекопитающих, а их оперение никогда не тускнеет. Имея физиологию, отличную от млекопитающих, они получили заметные эволюционные преимущества. Одно из них — уникальное зрение, совмещающее в себе навигацию по магнитному полю, невероятную остроту и восприятие ультрафиолета.

Конкурс «Био/Мол/Текст»-2021/2022
Эта работа опубликована в номинации «Свободная тема» конкурса «Био/Мол/Текст»-2021/2022.
Партнер номинации — компания SkyGen: передовой дистрибьютор продукции для life science на российском рынке.
Генеральный партнер конкурса — международная инновационная биотехнологическая компания BIOCAD.
Генеральный партнер конкурса — компания «Диаэм»: крупнейший поставщик оборудования, реагентов и расходных материалов для биологических исследований и производств.
«Книжный» спонсор конкурса — «Альпина нон-фикшн»
«Видимый» диапазон света. Так ли было всегда?
Физиология зрения и физика света тесно друг с другом связаны. В 1927 году академик С.И. Вавилов заканчивает свою книгу «Глаз и Солнце» словами: «Глаз нельзя понять, не зная Солнца. Вот почему глаз солнечен, по словам поэта» [2].
В чем же «солнечность» глаза? Для того, чтобы понять это, достаточно взглянуть на электромагнитный спектр излучения Солнца (рис. 1, сверху). Именно благодаря спектру испускания Солнца сформировались спектральные характеристики светочувствительных зрительных пигментов — палочек и колбочек, запускающих процесс зрения. На самом деле, человеческий глаз способен различить не весь спектральный диапазон, доходящий до поверхности Земли (рис. 1, снизу), но именно та его часть, которую могут распознать наши зрительные пигменты, названа «видимым светом» — 380–760 нм [3].

Рисунок 1. Электромагнитный спектр солнечного излучения. У поверхности Земли около 80% солнечной энергии ограничено спектральным диапазоном 300–1100 нм [3]. При этом «видимым светом» принято обозначать только ту область, которую способен распознавать человеческий глаз.
В ходе биологической эволюции зрительная клетка достигла предела чувствительности, допускаемой квантовой теорией света. Согласно данным физиков из Университета Иллинойса, человеку достаточно всего трех квантов света, чтобы увидеть вспышку, а отдельные клетки сетчатки (палочки) могут регистрировать и единичные кванты света. Однако вероятность, что сигнал, порождаемый только одной клеткой, дойдет до мозга, крайне мала [3]. Получается цепочка: один квант → поглотившая его молекула зрительного пигмента → одна возбужденная клетка сетчатки (палочка) → зрительный сигнал. Но чтобы распознать еле заметную вспышку света, одного кванта недостаточно. Обычно человеку требуется около 10–20 квантов света; тогда сигналы от 10–20 палочек суммируются и передаются в мозг, в его зрительные центры. Благодаря этому механизму человек способен различать на ночном небе очень слабые звездочки — шестой, а в некоторых случаях даже седьмой и восьмой звездной величины. Доходящий от звезд до Земли свет — это всего лишь 10−12—10−14 от полного солнечного света. Таким образом, наша зрительная система (глаза и мозг) может эффективно работать в огромном диапазоне освещенностей — от почти полной темноты до яркого солнечного дня.
Глаз животных и человека как нельзя лучше приспособлен для восприятия именно естественного солнечного света, который и сформировал «под себя» органы зрения всех животных — от самых примитивных беспозвоночных до высших позвоночных.
Фотоэлектронный умножитель глаза
Но как возможно, чтобы всего один квант света вызывал бурную реакцию в нашем глазу? «Биология становится слишком серьезной наукой, чтобы ее можно было доверять биологам», — пошутил кто-то из физиков. И оказался абсолютно прав!
Чтобы понять, как работает колбочка, надо разобраться в устройстве такого прибора, как фотоэлектронный умножитель (ФЭУ). Принципиальная схема его работы представлена на рисунке 3А (см. также врезку) и заключается в каскадном последовательном выбивании электронов с фотопластинок для увеличения количества электронов, попадающих на детектор. В случае с глазом, в роли детектора выступает нервное окончание.
Однако количество электронов, которые возможно выбить из пластинки динода, не бесконечно. Поэтому когда ФЭУ приходят в негодность, их меняют. А что же с глазом? Если посмотреть на схему строения колбочки глаза (рис. 2Б), то мы увидим, что ее наружный сегмент состоит из плазматической мембраны, наполненной стопками дисков с молекулами светочувствительных белков — родопсинов. В палочке квант света поглощается одной из 109 молекул родопсина. В результате чрезвычайно быстрой (менее 1 пикосекунды — 10−12 с) фотохимической реакции и затем последовательности темновых реакций, зависимых от температуры превращений, молекула родопсина обесцвечивается. Это означает, что каким-то образом в палочке эта одна обесцвеченная из 109 необесцвеченных молекул должна быть «узнана». Далее эта «узнанная» молекула запускает мощный каскад усиления первичного светового, фотохимического события, вследствие чего клетка отвечает миниатюрным, хотя и вполне ощутимым электрическим рецепторным сигналом. В последние годы удалось напрямую зарегистрировать этот миниатюрный сигнал.
Миниатюрный электрический ответ в палочке на один квант все же достаточно велик: он составляет около 3% от ее максимального ответа на яркую, насыщенную вспышку света. Такой величины сигнал может быть уверенно передан дальше — нейронам сетчатки, а затем в мозг.
Когда молекула родопсина «изживает» себя, диск за ненадобностью выкидывается из палочки, но так никуда и не исчезает из глаза. Именно поэтому у людей преклонного возраста белок глаза имеет желтоватый оттенок — это не что иное, как накопившиеся израсходованные диски с родопсином [3], [7].
Родопсин и фотоизомеризация. Молекула, благодаря которой мы видим свет
«Биомолекула» уже подробно рассказывала об этой замечательной молекуле, и прежде всего мы советуем читателям обратиться к статье «Зрительный родопсин — рецептор, реагирующий на свет» [8]. Являясь типичным представителем G-белок сопряженных рецепторов (подробнее о G-белках читайте в статье «Биомолекулы» «Аллостерические регуляторы GPCR: ключи от всех замков» [9]), родопсин переходит из неактивной в активную форму при поглощении кванта света ретиналем (рис. 3): главной реакцией этого фотопревращения является переход хромофорной группы родопсина — 11-cis-ретиналя — в all-trans-ретиналь.

Рисунок 3А. Схема фотопревращений родопсина. Промежуточные стадии фотопревращения родопсина при попадании кванта света. При этом конформационные перестройки белковой части молекулы происходят сначала в хромофорном центре, а затем и в цитоплазматическом домене. Родопсин переходит в активированное состояние и взаимодействует с G-белком трансдуцином (подробнее — в статье «Зрительный родопсин — рецептор, реагирующий на свет» [8]).

Рисунок 3Б. Молекула 11-cis- и all-trans-ретиналя.
Птичье превосходство
Зрение птиц отличается от человеческого и продиктовано их образом жизни. Обитание в воздушной среде, высокая маневренность и широкий обзор горизонта при наборе высоты способствовали максимальному развитию зрения у этого класса животных. Формированию детального изображения при высокой остроте способствуют крупные глаза, совершенная аккомодация хрусталика с помощью системы поперечнополосатых цилиарных мышц, возможность дополнительно изменить кривизну роговицы, васкуляризованный гребешок внутри глаза и сложно устроенная сетчатка с одной или двумя фовекальными ямками, не имеющая аналогов у других классов животных. До 1980-х годов изучению глаз птиц не уделяли особенного внимания, поэтому известно далеко не так много, как хотелось бы. Важный вклад в изучение глаз птиц внес Боумейкер с коллегами, изучив спектры поглощения зрительных пигментов фоторецепторов птиц [10]. Активное изучение механизмов зрения птиц началось в 2000-х годах. На сегодня известно, что обработку зрительного сигнала обеспечивают палочки и колбочки, но их свойства, морфология и биохимия отличны от человеческих и малоизучены. Механизм цветовосприятия у птиц на сегодняшний день так и не установлен.
Подробнее о том, как видят другие животные, можно прочитать в статьях «Биомолекулы» «Разные глаза — разные возможности» [11] и «Откуда пошло зрение» [12].
Вижу на все 360o (кроме кончика клюва)
Зрение — основное чувство дальней и ближней ориентации птиц. В отличие от других позвоночных, среди них нет ни одного вида с редуцированными или слепыми глазами. У птиц, ведущих ночной образ жизни, можно наблюдать трубчатые глаза с небольшим количеством детекторов цвета, но с многочисленными палочками, которые хорошо функционируют при слабом освещении. Аккомодация глаза достигается изменением формы хрусталика и его перемещением [13].
На рисунке 4 хорошо видно, что глаза птиц расположены таким образом, чтобы обеспечивать хорошее бинокулярное зрение. Различное расположение глаз (рис. 4Б, 4В) связано с особенностями жизни каждого вида. Так, сове, будучи ночным хищником, вовсе не обязательно видеть то, что творится у нее за спиной, в то время как цапля или вальдшнеп, постоянно пребывающие в поиске еды, имеют полный обзор вокруг своей головы. Все это сводит к минимуму возможность незаметно подкрасться к птице сзади. В то же время, если птица будет полностью сконцентрирована на изучении объекта вдалеке, она может не заметить подкрадывающегося хищника. Подробнее об этом мы поговорим в других разделах.
Если взглянуть на рисунок 4Б, то видно, что у вальдшнепа есть слепая зона на самом затылке и кончике клюва. Аналогично — у ястреба, воробья и цапли. Вот так: получается, что несмотря на широкий угол зрения, птица почти никогда не видит, что расположено у нее прямо под клювом.

Рисунок 4А. Поле зрения у разных видов птиц и других животных. На этой панели показано поле зрения ястреба с указанием зон моно- и бинокулярного зрения. Продолжение в крутилке!

Рисунок 4Б. Сравнение зон монокулярного и бинокулярного зрения у птиц разных видов.

Рисунок 4В. Сравнение зон монокулярного и бинокулярного зрения у птиц разных видов.
адаптировано из Medical Illustration & Animation

Рисунок 4Г. Поля зрения у зайца, совы и человека. У хищников больше развито бинокулярное зрение.
Как же они видят?
Глазное яблоко птиц — сложный рецепторный аппарат, основные структуры которого сходны со структурами глаз других позвоночных (рис. 6). Орган зрения состоит из глазного яблока со зрительным нервом и придаточных органов: век, слезных желез и двигательных мышц. Глазное яблоко спереди выпуклое, в задней части округлое, в середине в виде усеченного конуса. Яблоко имеет три оболочки — наружную, среднюю и внутреннюю. Сосудистая (средняя) оболочка находится под склерой; она состоит из собственно сосудистой оболочки, радужной оболочки и ресничного тела.
К центру сетчатки расположена центральная ямка с большой плотностью рецепторов. Это область наибольшей остроты переднего зрения, то есть резкого и четкого обнаружения объектов. У 54% птиц есть вторая центральная ямка для улучшенного бокового обзора. На оптическом диске у птиц есть небольшое слепое пятно, лишенное фоторецепторов; это место присоединения зрительного нерва и сосудов к глазу.
Птичий глаз напоминает глаз рептилии: он оснащен цилиарной мышцей, способной менять форму хрусталика быстрее и сильнее, чем у млекопитающих. В животном царстве у птиц самые большие глаза относительно размера тела. По внутреннему строению они схожи с глазами других позвоночных, но имеют уникальную структуру — гребень. Глаза птиц защищены третьей прозрачной подвижной мембраной.

Рисунок 5. Глаз ночной хищной птицы. Глазное яблоко птиц имеет уплощенную форму и малоподвижно. Однако этот «недостаток» природа компенсировала способностью удерживать в фокусе большое поле зрения. Другим подарком природы стал гребешок (цифра 6), обеспечивающий острое зрение. Глазной гребень отсутствует у млекопитающих и слабо развит у рептилий. Первое описание гребня датируется 1676 годом, и до сих пор этот орган остается предметом многочисленных исследований; и в то же время он слабо изучен. Его основная функция остается невыясненной; доподлинно известно только, что его формы и размеры у разных птиц различна. Основная функция гребня — снабжение стекловидного тела и внутренних слоев сетчатки кислородом и удаление продуктов метаболизма.
Гребень (или гребешок) — малоизученный орган складчатой ткани, который находится под сетчаткой. Он хорошо снабжен кровеносными сосудами и, вероятно, питает сетчатку, а также защищает ее от ослепительного света или помогает обнаруживать движущиеся объекты. Глазной гребень обильно наполнен гранулами меланина, которые поглощают рассеянный свет, попадающий в глаза птицы, уменьшая фоновые блики. Небольшое потепление гребня вследствие поглощения света гранулами меланина предположительно повышает интенсивность метаболизма гребня, увеличивая секрецию питательных веществ в стекловидное тело. Вероятно, очень высокая ферментативная активность щелочной фосфатазы в глазном гребне поддерживает его высокую секреторную функцию для дополнительного питания сетчатки. Складки гребня сетчатки глаза птиц представляют собой богатую сосудистую сеть. Интенсивный транспортный обмен метаболитами между кровью, внутриглазной жидкостью и структурами гребня сетчатки происходит путем активной диффузии [14–16].
Палочки, колбочки, а вы кто?
Сетчатка птиц имеет три типа фоторецепторов:
- Палочки обеспечивают черно-белое зрение при низкой освещенности.
- Колбочки — цветное зрение при ярком свете.
- Двойные колбочки — цветное зрение.
Птицы являются тетрахроматами и воспринимают ультрафиолет (рис. 7). Помимо этого, сетчатка птиц не имеет собственных сосудов, и это уменьшает аберрации высшего порядка, которые снижают качество зрения человека. Острое зрение птиц достигается за счет большого количества фоторецепторов. Для сравнения, максимально достижимая острота зрения зависит от плотности фоторецепторов на сетчатке глаза: у человека их количество составляет около 200 000 на 1 мм2, а у домашнего воробья — 400 000 на 1 мм2! Помимо этого, хищные птицы и птицы, способные развивать высокую скорость полета (ласточки) или «летать на месте» (колибри, зимородок), имеют вторую ямку для улучшенного бокового обзора. Другое главное преимущество птиц заключается в быстрой обработке информации во время полета. Чтобы «думать в полете», скорость восприятия зрения составляет от 130 до 150 Гц, в то время, как у человека — только 50 Гц [17]. С другой стороны, для «одомашненных» птиц подобная скорость восприятия света будет скорее помехой, чем преимуществом, ведь частота мерцания люминесцентной лампы составляет 50–60 Гц, поэтому для человека ее свет кажется непрерывным. А теперь представьте, как чувствует себя птица!

Рисунок 7. Цветовое восприятие птичьего зрения. Схематически пространство цветового зрения человека можно изобразить в виде области плоского треугольника (слева). На треугольнике отложены длины волн, воспринимаемых каждым из типов колбочек. Чтобы отразить цветовое зрение птицы, необходимо добавить еще одну ось к треугольнику (справа), которая будет отражать наложение от ультрафиолета на видимый спектральный диапазон. В результате получится тетраэдр: цвета в присутствии УФ отмечены плавающими шарами над основанием тетраэдра. Чем выше доля УФ в спектре, тем выше расположены точки. Например, оперение расписного овсянкового кардинала (внизу слева) отражает различное количество ультрафиолета в дополнение к цветам, различимых для человеческого глаза. Внизу справа изображен спектр отражения его оперения в видимом и УФ-спектре. Именно в УФ-области будут заметны различия по рисунку оперения.
Так же, как и у млекопитающих, у дневных видов птиц палочек всего один вид, но целых шесть видов колбочек (для сравнения: у человека и приматов — три вида, а других млекопитающих — два). Четыре вида колбочек, максимально чувствительных к фиолетовой (ультрафиолетовой), синей, зеленой и красной областям спектра, обеспечивают птицам богатое цветное зрение (рис. 7). Оставшиеся два вида соединены вместе и функционируют как единый фоторецептор. Они называются двойными колбочками, и их роль состоит в восприятии не цвета, а движущихся объектов. Количество разных колбочек не одинаково. В сетчатке бóльшую долю среди фоторецепторов занимают двойные колбочки (40,7%), затем зеленые (21,1%), красные (17,1%), синие (12,6%) и фиолетовые (8,5%) (рис. 8).
У птиц двойные колбочки представляют собой две плотно сросшиеся колбочки неравного размера. Их функция, по сравнению с обычными колбочками, до сих пор неясна, но главным образом они задействованы в ахроматических задачах — то есть не связаны с восприятием цветового зрения, а отвечают за регистрацию движения, определение яркости и поляризацию [10].
Встроенный светофильтр
Колбочки птиц, рыб и других позвоночных сохранили некоторые особенности, утерянные млекопитающими. Из них самое важное для цветового зрения — это наличие цветных капель жира. В зависимости от их типа, колбочки содержат красные, желтые, почти бесцветные и прозрачные липидные капельки. Цвет липидной капли определяется содержанием липидов, каротиноидов и специальным компонентом, образуя вместе природный светофильтр разного размера (рис. 9). При прохождении через липидную каплю отсекается часть длин волн, на которые реагируют колбочки, благодаря чему птицы способны различать большее количество цветов. Предполагается, что липидная капля образуется в результате жирового перерождения митохондрии во время дискогенеза или немного позже, после чего происходит их слияние и образование капли перед базальными дисками [10].

Рисунок 9А. Расположение колбочек с липидными каплями в сетчатке цыпленка. На микрофотографии сетчатки гаички хорошо заметны бесцветные, желтые и красные пятна. Все окрашенные капли служат светофильтрами, не пропускающими короткие длины волн. Зрительные пигменты SWS1 (воспринимают ультрафиолет) содержат самые маленькие, бесцветные капельки T (transparent) и обладают максимумом поглощения при 400 нм. Эти капли обладают голубовато-белой флуоресценцией в ультрафиолетовом свете (второе изображение слева). Размер самых маленьких капель составляет 1–2 мкм, а самых больших — около 5 мкм. Доподлинно известно, что цвет липидной капли может меняться с возрастом птицы, однако причины этого явления еще неясны [10], [21].

Рисунок 9Б. Благодаря липидным каплям птицы видят и различают гораздо большее количество оттенков цветов — по сравнению с человеком. В шуточной форме это можно изобразить в виде незамысловатой картинки.
коллаж автора
Организованный хаос
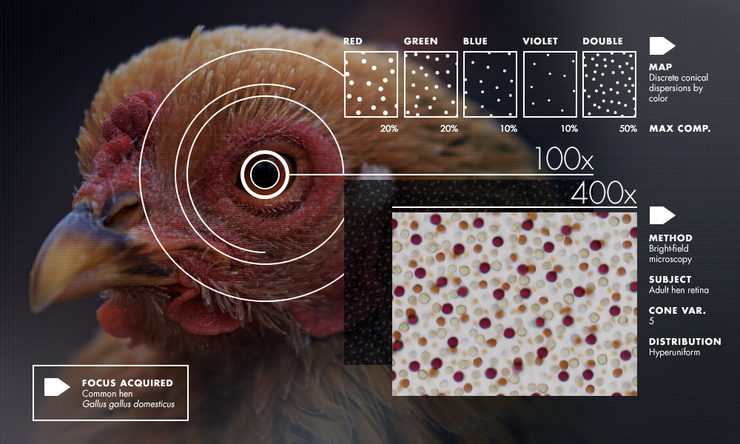
Большинство опытов по изучению глаз птиц проводится на цыплятах или взрослых курицах. Колбочки расположены на сетчатке неравномерно, неупорядоченно, но и не хаотично. Относительно друг друга они выстраиваются в сложную мозаику, причем вокруг колбочки одного цвета могут находиться колбочки только другого (но не того же самого) цвета. То есть, при наличии в глазу шести видов колбочек, сетчатку можно разбить на шесть видов сегментов мозаик, чтобы увидеть упорядоченность (рис. 10). Закономерность, найденная в сетчатке цыплят, оказалась верной и для других птиц [22].

Рисунок 10. Организованный хаос. Цветные точки в квадрате слева соответствуют расположению зеленых, синих, красных, фиолетовых и двойных (обозначено черным) фоторецепторов сетчатки курицы. На первый взгляд кажется, что колбочки перемешаны между собой беспорядочно, но на самом деле они имеют упорядоченность относительно колбочек своего вида, независимо от колбочек другого цвета. Таким образом, колбочка формирует вокруг себя «зону исключения» для колбочки своего же вида (обозначено закрашенными кружками на рисунках справа). При этом в «зоне исключения» могут располагаться колбочки всех других типов. Это распределение подчинено пространственному разделению. Каждый набор колбочек нельзя назвать идеально однородным, но он однороден настолько, что может иметь ограничения по распределению пяти различных колбочек. Гиперупорядоченость, в свою очередь, образует гипероднородность — то есть, если разбить сетчатку на сегменты условного размера, то количество всех рецепторов внутри каждого сегмента не будет сильно различаться.
Мигательная перепонка
Еще одно преимущество птиц, которое они сохранили (в отличие от млекопитающих) в процессе эволюции, — мигательная перепонка (рис. 11). Она представляет собой (полу)прозрачное третье веко, существующее у некоторых животных и позволяющее защищать глаз или увлажнять его без потери видимости. У человека она осталась в виде рудимента — третьего века в уголке глаза.

Рисунок 11. Мигательная перепонка (третье веко) у птиц и человека.
Вы не подскажете, как лететь на юг?
Некоторые виды птиц, в зависимости от времени года, совершают миграции к местам зимовок и обратно. Как им удается точно распознавать, где север, без карманного компаса? Помимо способности определять положение Солнца и звезд [24], природа снабдила перелетных птиц уникальным приспособлением, расположенным прямо в их глазу. Детали каждого из устройств не до конца ясны, но больше всего внимания ученые уделяют магнитному компасу, который по праву занимает главенствующую позицию среди имеющихся у птиц средств навигации. Следом за ним следует солнечный компас, принцип действия которого основан на чувствительности к поляризованному свету [25].
Некоторые виды птиц способны использовать для вычисления направления магнитного поля Земли квантовый компьютер, встроенный прямо в их глаза. Но прежде всего, стоит понимать, что птичий компас отличен от человеческого устройства — если привычный нам прибор указывает стрелкой на северный и южный магнитные полюса, то птичий навигатор будет определять положение «магнитного полюса» и «магнитного экватора». На самом деле, птицы реагируют на магнитное наклонение — угол, образуемый силовыми линиями поля с плоскостью горизонта, и им совершенно безразлично, где север, а где юг. Другая важная составляющая магнитного компаса птиц — освещение. Птичий навигатор полностью отказывается работать в темноте: ему требуется, пусть и слабое, освещение с длиной волны 400–570 нм. Более подробно о системе навигации пернатых можно прочитать в статье «Биомолекулы» «Пазл магнитного компаса птиц» [26].
Большинство исследователей полагает, что перелетные птицы способны воспринимать магнитное поле Земли благодаря процессам магниторецепции криптохрома. Птицы поворачивают голову, чтобы определить магнитное поле. В глазу перелетной птицы расположены чувствительные к свету белки криптохромы со светособирающим хромоформом птерином. Криптохром-4 выглядит самым подходящим претендентом на роль руководителя компаса. Во-первых, он экспрессируется сезонно (весной и осенью), а во-вторых, он расположен не везде, а только в красных и двойных колбочках.
Три консервативных аминокислотных остатка триптофана (Trp 400, Trp 377 и Trp 324; рис. 12) криптохрома являются ключевыми элементами: под действием синего света происходит перераспределение заряда между этой триадой и флавином. В результате образуется устойчивая радикальная пара, чувствительная к магнитному полю. Но даже это простое объяснение вызывает ряд вопросов, ведь криптохром поглощает в диапазоне 400–500 нм [27], в то время как птицы продолжают безошибочно ориентироваться и при более длинноволновом освещении.
Более подробно о магниторецепции и работе криптохрома можно прочитать в статье «Биомолекулы» «Нобель vs Шнобель, или Механизмы магниторецепции» [28].
Чувствительность птиц к изменениям магнитного поля весьма высока. Так, в экспериментах с европейскими малиновками изменение магнитного поля на 15 нТл (менее одной тысячной силы магнитного поля Земли) оказалось достаточным, чтобы нарушить чувство направления птицы [30]. Другое исследование показало, что состояние квантовой запутанности в глазах птицы должно длиться как минимум 100 мкс, чтобы птица чувствовала направление магнитного поля. На настоящее время ученым удалось создать установку, поддерживающую квантовую запутанность только 80 мкс, а птицы могут поддерживать это состояние благодаря затухающему осцилляторному колебанию электронов при их встрече с клетками глаза [31]!
Согласно гипотезе магниторецепции, молекулы криптохрома способны провзаимодействовать только с электронами с противоположным спином. Таким образом, если спины останутся в запутанном состоянии, это приведет к получению глазом энергии от фотона. Но если птица ориентирована «неправильно», то спины не становятся параллельными, не рекомбинируют, и оптический нерв не получит возбуждающего стимула. Согласно другому представлению, свет возбуждает криптохромы, за счет чего происходит испускание непарных электронов, взаимодействующих с магнитным полем Земли. То есть, существуют магниточувствительные бирадикальные химические реакции, которые чувствительны к расположению птицы в магнитном поле Земли, или, иными словами — чувствительны к углу между силовыми линиями магнитного поля и поверхностью Земли [32–35].
В настоящее время о зрении птиц известно очень мало, но в то же время, уже известные факты ставят ученых в тупик, а решение поставленных задач может служить толчком к развитию новых технологий!
Литература
- Зрительный родопсин — рецептор, реагирующий на свет;
- Вавилов С. И. Глаз и солнце. М.: «Академия наук СССР», 1927. — 334 с.;
- Островский М. А., Фельдман Т. Б. Фотобиология и фотохимия первичных процессов зрения. Дубна : Междунар. ун-т природы, о-ва и человека «Дубна», 2011. — 58 с.;
- Как исследовать клетку на уровне отдельных биомолекул;
- 12 методов в картинках: микроскопия;
- Крупные подробности микроскопического мира: Нобелевская премия по химии 2017;
- Островский М. А. (2005). Молекулярные механизмы повреждающего действия света на структуры глаза и системы защиты от такого повреждения. Успехи биологической химии. 45, 173–204;
- Зрительный родопсин — рецептор, реагирующий на свет;
- Аллостерические регуляторы GPCR: ключи от всех замков;
- Хохлова Т.В. (2009). Фоторецепторы птиц: молекулярная генетика зрительных пигментов, структурные и функциональные особенности клеток и их топография. Сенсорные системы. 23, 91–105;
- Разные глаза — разные возможности;
- Откуда пошло зрение;
- Terry M. Tritt, M. A. Subramanian. (2006). Thermoelectric Materials, Phenomena, and Applications: A Bird's Eye View. MRS Bull.. 31, 188-198;
- Шляпникова А. А. (2012). Морфологические особенности сетчатки глаза у птиц (куры, утки, гуси). Ученые записки Казанской государственной академии ветеринарной медицины им. Н.Э Баумана. 1, 212–216;
- Дегтярев В. В., Шляпникова А. А. (2015). Результаты электронно-микроскопических исследований ультраструктуры гребня сетчатки глаза птиц. Современные проблемы науки и образования. 3, 604–605;
- Соломахина Л. А. (2016). Нормальные вариации глазного дна птиц. VetPharma. 3, 56–65;
- Jannika E. Boström, Marina Dimitrova, Cindy Canton, Olle Håstad, Anna Qvarnström, Anders Ödeen. (2016). Ultra-Rapid Vision in Birds. PLoS ONE. 11, e0151099;
- Разные глаза — разные возможности;
- Lukas Hofmann, Krzysztof Palczewski. (2015). Advances in understanding the molecular basis of the first steps in color vision. Progress in Retinal and Eye Research. 49, 46-66;
- Evtukh G. (2019). Fluorescence among Fraterculinae subfamily. Русский орнитологический журнал. 28, 2134–2142;
- N. S. Hart, J. C. Partridge, I. C. Cuthill, A. T. D. Bennett. (2000). Visual pigments, oil droplets, ocular media and cone photoreceptor distribution in two species of passerine bird: the blue tit ( Parus caeruleus L.) and the blackbird ( Turdus merula L.). Journal of Comparative Physiology A: Sensory, Neural, and Behavioral Physiology. 186, 375-387;
- Yang Jiao, Timothy Lau, Haralampos Hatzikirou, Michael Meyer-Hermann, Joseph C. Corbo, Salvatore Torquato. (2014). Avian photoreceptor patterns represent a disordered hyperuniform solution to a multiscale packing problem. Phys. Rev. E. 89;
- Schwab I.R. (2002). Cure for a headache. Br J Ophthalmol. 86, 843;
- Н. С. Чернецов, N. S. Chernetsov. (2016). Ориентация и навигация мигрирующих птиц. Зоол. ж.. 95, 128-146;
- Зуева Л. В., Макаров В. И., Голубева Т. Б., Инюшин М. Ю. (2015). В поисках структурного базиса поляризационной чувствительности у птиц. Энергетика и годовые циклы птиц. 1, 126–130;
- Пазл магнитного компаса птиц;
- Л. А. Астахова, А. Ю. Ротов, К. В. Кавокин, Н. С. Чернецов, М. Л. Фирсов. (2019). Cвязь магнитного компаса и фоторецепции у птиц: гипотезы и нерешенные вопросы. Журн. общ. биологии. 80, 83-94;
- Нобель vs Шнобель, или Механизмы магниторецепции;
- PhilipR.A. May, Paul Newman, JoaquinM. Fuster, Ada Hirschman. (1976). WOODPECKERS AND HEAD INJURY. The Lancet. 307, 454-455;
- Thorsten Ritz, Roswitha Wiltschko, P.J. Hore, Christopher T. Rodgers, Katrin Stapput, et. al.. (2009). Magnetic Compass of Birds Is Based on a Molecule with Optimal Directional Sensitivity. Biophysical Journal. 96, 3451-3457;
- Erik M. Gauger, Elisabeth Rieper, John J. L. Morton, Simon C. Benjamin, Vlatko Vedral. (2011). Sustained Quantum Coherence and Entanglement in the Avian Compass. Phys. Rev. Lett.. 106;
- Кишкинёв Д. А.,Чернецов Н. С. (2014). Магниторецепторные системы у птиц: обзор современных исследований. Журнал общей биологии. 75, 104–123;
- Jingjing Xu, Lauren E. Jarocha, Tilo Zollitsch, Marcin Konowalczyk, Kevin B. Henbest, et. al.. (2021). Magnetic sensitivity of cryptochrome 4 from a migratory songbird. Nature. 594, 535-540;
- Norman Ginsburg, Virginia Nilsson. (1971). MEASURING FLICKER THRESHOLDS IN THE BUDGERIGAR1. Journal of the Experimental Analysis of Behavior. 15, 189-192;
- Henrik Mouritsen, Gesa Feenders, Miriam Liedvogel, Wiebke Kropp. (2004). Migratory Birds Use Head Scans to Detect the Direction of the Earth's Magnetic Field. Current Biology. 14, 1946-1949.



Комментарии
0Чтобы оставить комментарий, необходимо
войти