Вторая жизнь АТФ: от главной батарейки до нейромедиатора
11 января 2021
Вторая жизнь АТФ: от главной батарейки до нейромедиатора
- 9018
- 0
- 23
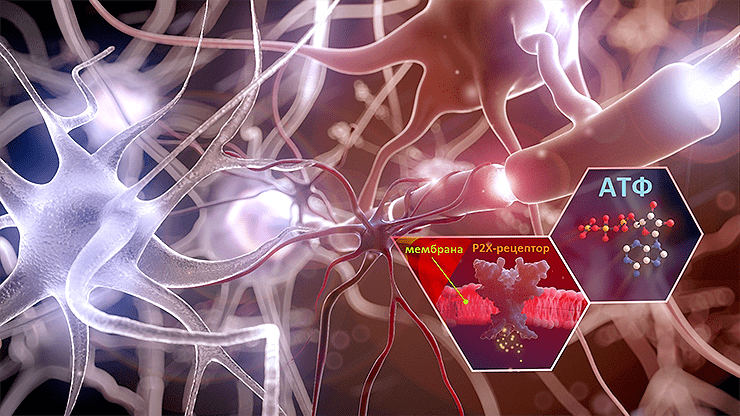
Микроглия, которая поддерживает работоспособность нервной системы, да и сами нейроны, вовсю экспонируют некие рецепторы. Активатором для них служит АТФ.
Longboat Partners with World-Leading Neuroscience Consortium, рисунок адаптирован
-
Автор
-
Редакторы
Темы
Статья на конкурс «Био/Мол/Текст»: Все знают АТФ. Но чем занимается АТФ вне клетки? Во что вылилась «пуримагическая гипотеза»? Каким образом клеточное «топливо», синтезируемое в организме буквально килограммами, становится тонким нейромедиатором? Какие клеточные рецепторы могут разрушать наш организм? Обо всём этом вы узнаете из данной статьи.

Конкурс «Био/Мол/Текст»-2020/2021
 Эта статья участвовала в конкурсе «Био/Мол/Текст»-2020/2021 в номинации «Свободная тема» и будет опубликована в журнале «Наука и жизнь».
Эта статья участвовала в конкурсе «Био/Мол/Текст»-2020/2021 в номинации «Свободная тема» и будет опубликована в журнале «Наука и жизнь».
Генеральный партнер конкурса — ежегодная биотехнологическая конференция BiotechClub, организованная международной инновационной биотехнологической компанией BIOCAD.

Спонсор конкурса — компания SkyGen: передовой дистрибьютор продукции для life science на российском рынке.
Спонсор конкурса — компания «Диаэм»: крупнейший поставщик оборудования, реагентов и расходных материалов для биологических исследований и производств.
«Книжный» спонсор конкурса — «Альпина нон-фикшн»
Вездесущий АТФ
Аденозинтрифосфат (АТФ; рис. 1) — это одна из самых важных молекул в нашем организме. Чаще всего АТФ рассматривается как универсальное топливо для многочисленных «молекулярных машин» и реакций в наших клетках. АТФ одна из самых реакционноспособных молекул на планете. Без него сама жизнь в том виде, в котором она есть сейчас, была бы невозможна.

Рисунок 1. Аденозинтрифосфат (АТФ) — «батарейка» всех клеточных процессов, состоящая из пуринового основания аденина, сахара рибозы и трех остатков фосфорной кислоты, которые хранят в своих связях внушительное количество потенциальной энергии.
Изначально его источником служил лишь бескислородный процесс гликолиза в цитоплазме. Такой способ оказался довольно неэффективным и мог снабжать энергией лишь доядерных прокариот. Появление в клетках верных вассалов-митохондрий позволило вывести производство АТФ на качественно новый уровень и получать его в недостижимых ранее количествах. В митохондриях это происходит при помощи цикла Кребса. Благодаря окислительному фосфорилированию, протекающему в этих органеллах, и клеточному дыханию возникло всё наблюдаемое нами многоклеточное биоразнообразие. Поэтому АТФ в клетках стало много, а за сутки общее количество расходуемого и вновь синтезируемого вещества в нашем теле исчисляется килограммами. Подробнее про синтез АТФ и появление митохондрий можно прочитать на «Биомолекуле» [1].
Более удивительным оказалось то, что и молекула АТФ, и «топливные отходы», получающиеся в результате разрыва фосфатных связей (энергия которых и используется клетками живых организмов для поддержания своей жизнедеятельности), являются частью обширного сигнального аппарата. Эта древняя система сигнализации охватывает весь организм и запускает сложные, порой необратимые, клеточные процессы. Она сообщает нам, когда идти спать, и она же заставляет нас кричать от боли. Находясь на службе клеточного иммунитета, она может как спасти организм от инфекции, так и привести к гибели клеток, став причиной множества воспалительных заболеваний. Более того, АТФ напрямую участвует в регуляции нашей нервной деятельности и служит эффективным нейромедиатором. Однако далеко не сразу эта функция стала очевидной. Для этого потребовались годы усердной работы и исключительное упрямство одного исследователя.
История одного открытия
Люди всегда будут критиковать. Прислушивайся к каждому возражению и проверяй его на практике. И если вдруг окажешься неправ, то ты должен быть первым, кто об этом объявит!
Началось всё с того, что в начале 1960-х годов британский ученый-нейробиолог Джеффри Бёрнсток (рис. 2) совершил открытие [2].

Рисунок 2. Джеффри Бёрнсток (1929–2020 гг.). Человек, который своими исследованиями подарил АТФ новую жизнь.
Пребывая в Австралии, Бёрнсток исследовал возбуждение гладкой мускулатуры кишечника морской свинки при помощи электрического тока. Можете не волноваться, он работал на срезе ткани, а не на живой свинке. В то время появились новые электрохимические методы, освоив которые, ученому не терпелось перейти к практике. Ему стало интересно проверить, как гладкая мускулатура будет реагировать на ток в условиях полной блокировки всех известных нервных рецепторов в конкретном срезе ткани. Частая практика у нейробиологов — заблокируй лишние рецепторы нейротоксинами, которыми нас снабдила сама эволюция в лице ядовитых растений и животных (ну и заодно химиков-синтетиков), а потом спокойно изучай то, что осталось рабочим.
Для начала он обработал ткань 6-гидроксидофамином (рис. 3а). Это синтетический нейротоксин, который нарушает работу дофаминовых и адренергических рецепторов. Затем нужно было заблокировать второй известный тип рецепторов — ацетилхолиновые. Для этого он накачал ткани атропином (рис. 3б), чтобы наверняка отключить все холинорецепторы.

Рисунок 3. Блокаторы Бёрнстока. а — 6-гидроксидофамин (он же 6-OHDA). б — Атропин — растительный алкалоид, известный блокатор ацетилхолиновых рецепторов.
Далее он ввел электрод и простимулировал ткани коротким электрическим импульсом. Ожидалось, что гладкая мускулатура неотвратимо сократится под действием электричества, однако ничего подобного не произошло. Какая-то неведомая сила противодействовала импульсам тока и запускала процессы, мешающие сокращению. На записях электрических сигналов наблюдалась выраженная гиперполяризация, а мышцы расслаблялись даже при серии таких импульсов.
На его счастье, в тот момент Бёрнсток работал с аспирантом из Японии, который использовал свои связи на родине и достал для исследователя знаменитый тетродотоксин (рис. 5а) из рыбы фугу (рис. 5б). Более подробно про тетродотоксин и с чем его едят, можно узнать, прочитав статью, опубликованную на «Биомолекуле» моим однокурсником: «Тетродотоксин — история элегантного убийцы» [4]. Вкратце, этот токсин известен тем, что может блокировать нервную проводимость и как пробка затыкать натриевые каналы. Однако он не лишает гладкую мускулатуру способности сокращаться. Смертоносное вещество было также принято Бёрнстоком в работу.

Рисунок 5а. Остолбеней! Структура тетродотоксина довольно уникальна по своей природе. Благодаря положительному заряду гуанидиновая группа ловко маскируется под гидратированные ионы натрия и проникает в пору натриевого канала. Однако за ней идет объемная клетка, сшитая из атомов углерода и кислорода, и утыканная гидроксильными группами. Она накрепко застревает в лоне ионного канала, препятствуя ходу ионов.

Рисунок 5б. Остолбеней! Рыба фугу — самый известный источник тетродотоксина. Однако виной всему не сама рыба, а обитающие в ней бактерии-симбионты. Именно они служат поставщиками яда в ее органах. Фугу считается экстремальным деликатесом в Японии, а для права ее приготовления повару требуется специальная лицензия. Остаточные дозы тетродотоксина в мясе все еще способны вызвать онемение губ и языка у отъявленных гурманов.
После его применения реакция на электрический разряд наконец стала такой, какой ожидалось — гиперполяризация пропала, а мышцы сократились. Вывод был очевиден: в нервах гладкой мускулатуры кишечника присутствовали неизвестные рецепторы (не холинергические и не адренергические), которые противодействовали сокращению и выполняли функции, тормозящие нервный импульс [5]. Начался усиленный поиск вещества, которое могло бы служить активатором этих новых загадочных рецепторов. К поиску применялся ряд стандартных критериев, таких как:
- нейромедиатор должен синтезироваться и храниться на нервных окончаниях, чтобы как можно быстрее передавать сигнал через синапсы (места контактов между двумя нейронами или нейроном и связанной с ним клеткой);
- должны существовать внеклеточные реакции, которые имитировали бы активацию таких рецепторов;
- высвобождаемый нейромедиатор должен тут же разрушаться какими-либо внеклеточными ферментами, чтобы им нельзя было «переборщить»;
- у таких рецепторов должны существовать свои блокаторы.
Однако испробованные нейропептиды, моноамины и аминокислоты не удовлетворяли заданным критериям. Как это часто бывает, подсказки лежали под ногами — следовало лишь немного покопать. Дело в том, что задолго до работы Джеффри Бёрнстока множество ученых уже натыкались на странное воздействие внеклеточных пуринов (гетероцикл пурина служит основой для аденозина в молекуле АТФ) на сердечную мышцу и кровеносные сосуды, но не придали этому большого значения. Например, знаменитый биохимик Альберт Сент-Дьердьи (рис. 6а) — тот самый, что впервые выделил витамин С (одновременно с Чарльзом Кингом), — также провел ряд фундаментальных исследований мышечного сокращения. В своей работе 1929 года Сент-Дьерди отметил, что введение пуриновых соединений в кровь животным влияло на их сердечный ритм [6]. Позже Памела Холтон (рис. 6б) в 1959 году замечала, что АТФ выделяется при стимуляции нервов в ушных артериях кролика и ведет к расслаблению стенок сосудов [7]. Однако никто из них не пошел в своих рассуждениях дальше и не узнал, насколько глубока эта кроличья нора.

Рисунок 6. Исследователи, работы которых помогли Джеффри Бёрнстоку отыскать новый нейромедиатор. Альберт Сент-Дьёрдьи (а) и Памела Холтон (б).
Вооружившись этими знаниями и проведя ряд опытов, Бёрнсток со своим коллегой Дэвидом Сэтчеллом доказали, что АТФ действительно активирует как стимулирующие, так и тормозящие эффекты в разных типах тканей организма. Статья об этом вышла в 1970 году [8]. Тем более, тогда уже стало известно о внеклеточных ферментах, способных расщеплять АТФ. Ничего не оставалось, кроме как признать, что аденозинтрифосфат является тем самым нейромедиатором и выдвинуть гипотезу о пуринергической передаче сигнала. Слово «пуринергическая» придумал сам Бёрнсток. Рецепторы, связанные с этим типом передачи сигнала, позже стали называть «пуринергическими», или «пуринорецепторами». Вернувшись в Англию, ученый столкнулся с недоверием со стороны коллег. В то время также был популярен так называемый принцип Дейла. Этот принцип Бёрнсток и многие другие нейробиологи того времени трактовали как утверждение, что один нейрон может осуществлять передачу сигнала при помощи только одного нейротрансмиттера (например ацетилхолина) и уж никак не при помощи нескольких разных. Тем более, при помощи АТФ. К слову, сам Генри Дейл имел в виду немного другое, но кто его слушал... Всем казалось маловероятным, что вездесущая молекула, источник энергии клеток, может участвовать в тонких процессах регуляции нервной деятельности.

Рисунок 7. Джеффри Бёрнсток. Фотография сделана в 1970 году в Мельбурне — как раз в тот год, когда была опубликована его первая статья о роли АТФ в качестве нейромедиатора.
В 1972 году вышла большая обзорная статья с выдвинутой ученым гипотезой о пуринергических нейротрансмиттерах и их возможной совместной трансмиссии с другими рецепторами нейронов (долой принцип Дейла!) [9]. Эта статья вызвала большой резонанс и стала причиной множества дебатов. Ее можно смело назвать кульминацией сделанных Бёрнстоком открытий в рамках его концепции о пуринергической передачи сигнала. В ней Бёрнсток также сетовал, что результаты его работ списывают на артефакты и ошибки эксперимента. Гипотеза Джеффри продолжала получать сопротивление в научных кругах на протяжении почти двух десятков лет. Некоторые оппоненты заявляли, что посвятят жизнь разрушению «пуринергической теории». Уважаемые ученые спорили до хрипоты, а один и вовсе назвал Бёрнстока «изобретателем пуримагической гипотезы» [2].
Но исследователь не сдавался и упрямо продолжал работу. Следующим шагом предстояло выяснить, что же собой представляют рецепторы, которые могут активироваться сигнальными молекулами АТФ, либо продуктами его распада (АТФ → АДФ → АМФ → аденозин → аденин).
В те времена, еще до бума генной инженерии, выделить и определить структуру этих рецепторов не представлялось возможным. Оставалось действовать наощупь — то есть скрупулезно копить данные и логически их сопоставлять. В 1978 году Бёрнстоку удалось выяснить опытным путем, что пуринергические рецепторы можно разделить на два семейства , которые он назвал P1 и P2 (P — пуриновые) [10].
Когда я говорю «семейство рецепторов», имеется в виду то, что раньше (в эволюционно древние времена) они кодировались одним геном, но в результате генетической дупликации возникли его видоизмененные копии, благодаря которым появилось разнообразие родственных рецепторов. Такие белки обычно объединяются в одно семейство и сохраняют высокое сходство последовательности и структурной укладки, но могут выполнять разные функции. Это разнообразие особенно велико, если ген белка — ключевой в выживании и эволюции вида при естественном отборе. Например, яды актиний, змей и пауков накопили целые комбинаторные библиотеки из многочисленных видоизмененных копий одного гена какого-нибудь пептидного токсина (который ранее вообще мог исполнять роль протеазного ингибитора, а не молекулярного оружия). Происходило это в результате эволюционного поиска и адаптации яда к своим жертвам, устойчивость которых тоже не стояла на месте в процессе эволюции. Подробнее про это читайте в статьях «Великому комбинатору и не снилось: комбинаторика токсинов пауков» [11] и «Яды — высокоточное оружие: компьютерное исследование природных нейротоксинов» [12]. Разнообразие же родственных пуринорецепторов у млекопитающих обусловливается усложнением организма и нервной системы в целом.
Это деление на семейства было условным и сделано исключительно по фармакологическим показателям. Рецепторы семейства P1 блокировались метилксантинами и активировались аденозином, в то время как члены семейства P2 в основном реагировали на АТФ, а метилксантины были над ними не властны.
В 1985 году Бёрнсток установил, что и P2-рецепторы можно поделить на два подсемейства — P2X и P2Y [13]. Было это сделано, опять же основываясь на их фармакологии и разной чувствительности к АТФ и его метиленовым производным.
Однако развитие методик молекулярного клонирования [14], [15] в начале 90-х наконец приоткрыло завесу тайны над загадочными пуринорецепторами. Вначале были клонированы и охарактеризованы рецепторы P1, а потом дошла очередь и до P2. А дальше завертелось. Бёрнстоку окончательно поверили. «Пуримагическая гипотеза» потеряла свою магию и перестала быть гипотезой. Исследователи со всего мира бросились изучать новые рецепторы, а количество публикаций на тему «пуринергическая передача сигнала» начало расти буквально в геометрической прогрессии (рис. 8).

Рисунок 8. Количество работ, посвященных пуринергической передаче сигнала, в период с 1972 по 2012 год. Как видно из графика, начало 90-х стало переломным моментом в этой сфере исследований.
Также пуринергическую передачу сигнала стали называть «вторая жизнь», или «вторая профессия АТФ». Таким образом, Джеффри Бёрнсток оказался на коне и моментально попал в топ самых цитируемых ученых в мире. Благодаря развитию рентгеновской кристаллографии и электронной микроскопии, с конца нулевых и на протяжении 2010-х были получены трехмерные структуры рецепторов, позволившие нам разглядеть их строение. Еще одно фундаментальное знание о функционировании нашего (да и не только нашего) организма легло в копилку современной биомедицины.
Пуринергическая передача сигнала: что мы знаем теперь?
А знаем мы довольно много. Пуринергическая передача сигнала отвечает не только за нейромедиацию. Это комплексная система, затрагивающая все уровни организма. Пуринорецепторы отвечают за осязание, боль, регуляцию сердечного ритма, сокращение гладкой мускулатуры, а также выделение желчи и эндокринных гормонов. Что немаловажно, они вовлечены в клеточный иммунитет, а нарушение их работы служит причиной разных патологических состояний в организме. «Принцип Дейла» к нашему дню окончательно потерял актуальность. Теперь всем известно, что один нейрон может экспонировать на своих нервных окончаниях сразу несколько разных рецепторов и использовать для передачи сигналов разные молекулы-нейромедиаторы.
Пуринергические рецепторы работают в совокупности с другими нейронными рецепторами и регулируют нашу нервную деятельность. Классификация Джеффри Бёрнстока верна и работает до сих пор, за тем исключением, что в 2002 году у грызунов смогли найти и клонировать пуринорецепторы третьего семейства, которое реагирует на аденин — последнюю ступень деградации АТФ внеклеточными ферментами (казалось бы, ну сколько уже можно резать несчастную молекулу?) [16]. Новое семейство назвали P0. Оно насчитывает всего два представителя — AdeR1 и AdeR2 (то есть «адениновые рецепторы 1 и 2»), которые были обнаружены в центральной нервной системе лабораторных мышей, крыс и хомяков. Чтобы не запутаться в моем рассказе, лучше поглядывать на классификацию всех этих товарищей, представленную на рисунке 9.

Рисунок 9. Классификация пуринорецепторов
схема создана автором статьи при помощи Flowchart Maker & Online Diagram Software
Эта надежная, как швейцарские часы, система работает по цепочке (рис. 10). Сперва из клетки выделяется АТФ. Это происходит разными путями. Оказалось, что существуют специальные каналы и полуканалы (такие как паннексин-1) для высвобождения нуклеотидов, а также особые белки-переносчики для их переноса во внеклеточную среду. (Эволюция системы по переносу пуринов во внеклеточное пространство, пожалуй, заслуживает отдельной статьи.) Однако чрезвычайно много АТФ может выделиться, если содержащие его клетки будут повреждены или полностью разрушены в результате полученных ранений или некроза тканей. Это так называемый литический путь — то есть связанный с лизисом клеточной мембраны. На заре открытия пуринорецепторов такое выделение АТФ во внеклеточную среду казалось единственно возможным путем их активации. Сюда мы также запишем всевозможные травмы и переломы, в том числе хирургическое вмешательство. Всё это неизбежно приводит к чрезмерной активации пуринорецепторов и ко всяким нехорошим стрессовым состояниям. В целом, массовое выделение АТФ во внеклеточное пространство — это один из главных сигналов опасности для организма.

Рисунок 10. Упрощенная схема пуринергической передачи сигнала. Через специальные полуканалы, такие как паннексин-1, во внеклеточное пространство выходит АТФ, который, вкупе со своими продуктами распада, активирует разные пуринорецепторы.
[17], рисунок адаптирован
Естественно, АТФ может высвобождаться из везикул в синапсах, как и положено всем порядочным нейромедиаторам. Аденозинтрифосфат, будучи первичным передатчиком сигнала и нейромедиатором, в первую очередь воздействует на трансмембранные ионные каналы семейства P2X. Эти рецепторы мгновенно запускают потенциал действия по нейронам, а также способны активировать различные биохимические пути, благодаря току входящих в клетку ионов.
Далее вступают в дело различные семейства рецепторов, сопряженных с G-белком, или GPCR (от англ. G-protein coupled receptors). Первым подхватывает эстафету семейство P2Y — некоторые члены которого также активируются АТФ (рис. 11а). Затем внеклеточные ферменты, которые называются 5′-эктонуклеотидазами, по очереди отрезают от АТФ остатки фосфорной кислоты. Все продукты такого распада находят свое применение.
Первой ступенью деградации нуклеотида окажется аденозиндифосфат (АДФ) (рис. 11б). Он пойдет на активацию других членов подсемейства P2Y. Также некоторые «игреки» реагируют на целый пул различных пиримидинов с остатками фосфорной кислоты: УТФ (рис. 11в), УДФ (рис. 11г) и УДФ-глюкозу (рис. 11д).

Рисунок 11. Лиганды-активаторы пуриновых рецепторов P2Y. а — Аденозинтрифосфат (АТФ). б — Аденозиндифосфат (АДФ). в — Уридинтрифосфат (УТФ). г — Уридиндифосфат (УДФ). д — Уридиндифосфат-глюкоза (УДФ-глюкоза).
Затем АДФ распадется до аденозинмонофосфата (АМФ), а тот, в свою очередь, до аденозина (рис. 12а) — каркаса АТФ, лишенного остатков фосфорной кислоты. Аденозин окажет влияние на сопряженные с G-белком рецепторы семейства P1. К слову, аденозин может получаться при разрушении цАМФ («посыльного» G-белков), а также производиться в клетке de novo.
Однако на аденозине дело не заканчивается. У грызунов есть свои уникальные GPCR — вышеупомянутые рецепторы P0. В качестве лиганда они выбрали продукт распада аденозина — молекулу аденина (рис. 12б). P0-рецепторы изучены пока недостаточно хорошо, а существование их у людей до сих пор спорно.

Рисунок 12. Лиганды для P1- и P0-рецепторов. а — Аденозин — предпоследняя ступень деградации АТФ. Потерял фосфатные группы, но сохранил свой «сахарок» — рибозу. б — Аденин — голый остов от АТФ, лишенный фосфатных остатков и рибозы.
P2X: хороший, плохой, трансмембранный
Мы знаем множество историй, как дельфины помогали утопающим доплыть до берега. Просто некому было рассказать, как они толкают людей в противоположную сторону.

Рисунок 13. Ричард Алан Норт. Ученый, который работал во главе исследовательской группы, выделявшей и клонировавшей ДНК P2X-рецепторов с 1993 по 1998 годы.
P2X-рецепторы — это самые настоящие мембранные ионные каналы (ионотропные рецепторы), которые активируются АТФ и только АТФ. Исходя из названия статьи, на них мы остановимся более подробно. Хоть я и рассматриваю все семейства рецепторов в рамках пуринергической передачи сигнала, стоит оговориться, что P2X не имеет ничего общего с относительно родственными между собой GPCR типа P0, P1 и P2Y.
Семь разновидностей P2X-рецепторов найдены по всему организму млекопитающих, включая клетки нервной системы (рис. 10). Однако распространены они неравномерно, и разные клетки синтезируют разные наборы этих рецепторов. Тип клеток определяет состав набора и функции, которые в них выполняют рецепторы. Их ДНК была клонирована, а последовательности охарактеризованы на протяжении 1990-х. Здесь нужно отдать должное научной группе под руководством Ричарда Алана Норта (рис. 13).

Рисунок 14. Упрощенная схема одной из трех субъединиц P2X-рецептора. Каждая субъединица состоит из двух трансмембранных спиралей (ТМ1 и ТМ2). Концы внутри, а снаружи внеклеточная петля. При открытии канала подвижна TM2, а ТМ1 остается статичной.
В 2009 году наступил следующий важный этап в изучении P2X-рецепторов. На этот раз группа американских ученых под руководством Тосимицу Каватэ при помощи рентгеноструктурного анализа впервые получила трехмерную структуру рецептора P2X4 из рыбки Danio rerio [18] (спойлер: ученые на этом не остановились и продолжили получать структуры всех остальных «иксов» из разных организмов). Наконец-то мы увидели P2X-рецепторы воочию! Ионный канал оказался тримером, собранным из трех отдельных белковых молекул-субъединиц. Знатоки отметят, что похожей структурой тримера обладают кислоточувствительные ионные каналы (ASICs, от англ. acid-sensing ion channels), однако их аминокислотная последовательность не имеет ничего общего с P2X-рецепторами.
Аминокислотная последовательность субъединиц определяет тип P2X-рецептора. Каждая субъединица пронизывает клеточную мембрану двумя спиралями, ее N- и С-концы находятся внутри клетки, а над мембраной во внешний мир возвышается замысловатый домен (рис. 14).
Все три части, сложенные вместе, могут раскрываться и закрываться, подобно лепесткам на объективе фотоаппарата, формируя ионный канал. Но не это было самым замечательным. Каждая субъединица неуловимо напоминала всем образ дельфина (рис. 15а)! А весь рецептор целиком — это три переплетенных в объятиях млекопитающих (рис. 15б). Ученые до сих пор используют этот метафорический образ для того, чтобы свободнее ориентироваться в строении рецептора и рассуждать об аминокислотных остатках на спинном плавнике или, например, в голове. В целом, такое строение характерно для всех P2X-рецепторов.

Рисунок 15. Мне под кожу бы, под кожу мне / Запустить дельфинов стаю!.. а — Схематичное строение субъединицы P2X-рецептора в виде дельфина. б — Три таких «дельфина»-субъединицы, объединенные в рецептор-тример.
Единственным эндогенным ключом к дельфиньему братству является АТФ. Три молекулы этого нуклеотида встраиваются в карманы между «дельфинами» и производят открытие ионного канала (рис. 16). Все рецепторы, несмотря на одинаковую структурную укладку и тесные родственные связи, по-разному реагируют на АТФ.

Рисунок 16. Механизм работы P2X-рецептора. а — Закрытый P2X-рецептор (вид сбоку и сверху). б — Открытый рецептор. Для активации рецептора и открытия канала необходимо встраивание трех молекул АТФ (выделены зеленым) в консервативные сайты связывания.
Кому-то из P2X хватает маленькой концентрации АТФ, чтобы открыться и быстро схлопнуться обратно, на время потеряв чувствительность. Кто-то продолжает работать без остановки, пока рядом есть АТФ. Уникальный в своем роде P2X7-рецептор имеет несколько режимов работы: один для низких и один — для высоких концентраций агониста. При продолжительном воздействии большого количества АТФ он раскрывается еще шире, образуя гигантскую пору в клеточной мембране. От таких огромных порталов во внешний мир клетка явно будет не в восторге.
- Агонист
- активатор рецептора. В противоположность антагонисту, который мешает его работе.

Рисунок 17. Классификация P2X-рецепторов и их гетеромеров. Выделенный темно-серым P2X6-тип не может формировать рецептор без помощи других типов, а вот у P2X7 (красный), наоборот, отсутствуют гибриды с другими типами рецепторов (пока спорно).
схема, создана автором статьи при помощи Flowchart Maker & Online Diagram Software.
Стоит отдельно сказать о существовании в организме «химер», которые слеплены из субъединиц различных типов и комбинируют их свойства (рис. 17). Например, субъединица P2X6 может существовать только в виде микса с субъединицами других типов. А вот уникальный P2X7 настолько отличается от других, что формирует рецептор исключительно из своих субъединиц [21].
P2X-рецепторы — это неселективные ионные каналы, которые не специализируются на каком-то конкретном типе ионов. Это весьма разносторонние личности. Они пропускают ионы калия, натрия, кальция, а иногда и хлора. А еще через них, как песок сквозь пальцы, калий может вытекать наружу. Однако именно способность пропускать кальций делает их такими многофункциональными. Ведь ионы кальция — это важнейший передатчик внутриклеточных сигналов в организме (один из вторичных мессенджеров), от него зависят многие каскады реакций. Наше сердце бьется в такт благодаря строго выверенным дозам кальция и не без помощи P2X-рецепторов [22]. Не зря кальцию посвящен целый научный журнал под названием Cell Calcium. Никакие другие ионы такой чести не удостаивались. Как тебе такое, натрий?
P2X-рецепторы выполняют самые разнообразные функции в организме, а каждое новое исследование открывает целый пласт процессов, в которые они оказываются вовлечены. Часто, благодаря их схожести, роли «иксов» пересекаются друг с другом, в зависимости от их локализации. Главными из них принято считать регуляцию сердечно-сосудистой деятельности и передачу сигналов в нервной системе. Тем не менее для того, чтобы рассказать про каждый аспект их влияния, придется написать целую книгу, поэтому пробежимся по наиболее важным и интересным функциям.
Например, семявыводящие протоки у мужчин сокращаются во время эякуляции при помощи P2X1-рецептора. Мыши-самцы с нокаутированным геном этого рецептора не могли размножаться. Благодаря чему его даже рассматривали как возможную мишень для создания мужских противозачаточных средств [23], [24]. В кровеносной системе он опосредует сокращение сосудов в ответ на АТФ, который выделяется совместно с норадреналином в качестве нейромедиатора симпатических нервов [25].
P2X2 и его гибриды широко распространены в нейронах, где передают сигналы на синапсах и регулируют нашу нервную деятельность, воздействуя на другие нервные рецепторы, такие как GABA- [26] или NMDA-рецептор [27]. Также известно, что мыши с мутировавшим геном P2X2-рецептора теряли слух [28].
Активация P2X3-рецептора вызывает у нас сокращение мочевого пузыря при мочеиспускании. А еще он крайне необходим в работе вкусовых рецепторов, активации макрофагов, агрегации тромбоцитов и создании болевого сигнала [29].
Что касается последнего, то P2X3-рецептор выступает в роли проводника острой, ноцицептивной боли. К такой можно, например, отнести как боль от пореза пальца листом бумаги, так и боль, вызванную гибелью тканей в результате воспаления. Оказалось, что его эффективные блокаторы можно обнаружить в природе. В Институте биоорганической химии (ИБХ РАН) группа ученых под руководством академика Евгения Васильевича Гришина и при участии Александра Александровича Василевского открыла и выделила пептид из яда пауков рода Alopecosa. Этот пептид показал эффективное ингибирование тока ионов через P2X3-рецептор и был назван «пуротоксин-1» (то есть токсин, блокирующий пуриновые рецепторы) [30]. Новый токсин составил серьезную конкуренцию уже известным синтетическим низкомолекулярным блокаторам P2X-рецепторов. Он показал отличное обезболивающее действие в экспериментах на мышах. При этом его эффективная концентрация оказалась на три порядка меньше, чем у известного низкомолекулярного блокатора A-317491 (для достижения сопоставимого анальгетического эффекта). Пуротоксин-1 прошел все стадии доклинических испытаний и, вполне возможно, через несколько лет окажется на фармацевтическом рынке в качестве ультрасовременного анальгетика. Можно лишь отметить, что яды животных всё еще остаются удивительным источником уникальных биологически активных соединений, которых создали лучшие «биохимики» на этой планете — эволюция и естественный отбор.
P2X4-рецептор экспонируется на клетках эндотелия сосудов и приводит (в отличие от P2X1) к их релаксации. Таким образом он реагирует на АТФ, который высвобождается из эндотелия в условиях стресса при сжатии сосудов (как бы балансирует чаши весов, чтобы мы случайно не померли от гипертонии) [31]. Также он увеличивает сокращение сердечной мышцы [32]. В общем, P2X1 и P2X4 — это те рецепторы, воздействуя на активность которых мы сможем снизить риски сердечно-сосудистых заболеваний. Весьма перспективные мишени для разработки лекарств.
А еще рецепторы P2X4 и P2X7 собаку съели на клеточном иммунитете, воспалении и программируемой гибели клеток [33], [34]. Они активно экспонируются на большинстве иммунных клеток, таких как макрофаги, дендритные или тучные клетки. Без них не обходится ни одно инфекционное заболевание, травмы или вечеринки по поводу внезапных аллергических реакций. У них также самая высокая степень родства и сходства аминокислотных последовательностей [35]. Есть работы, которые говорят о возможности существования гетеромеров между ними на моделях in vitro [36], хотя пока это не было подтверждено в клетках реального организма.

Рисунок 18. Схема субъединицы P2X7-рецептора. Отличительной особенностью этого рецептора является длинная С-концевая цепочка, уходящая вглубь клетки.
Про P2X7-рецептор стоит поговорить отдельно. Ведь это целый кладезь сюрпризов. Попав впервые в центр внимания ученых в 1980 году, он здорово смутил исследователей. Его даже хотели вынести в новое отдельное подсемейство — P2Z [37]. Но, установив аминокислотную последовательность, немного успокоились и оставили в «иксах» [38] (хотя название P2Z нет-нет, да проскальзывает). Отличает его от прочих «дельфинчиков» более длинный С-конец, уходящий вглубь клетки. Это значительно увеличивает размер его субъединицы и существенным образом влияет на сигнальные свойства самого рецептора (рис. 18) [39].
Он обладает гораздо меньшей чувствительностью к АТФ, чем другие его собратья. Но как только агониста станет слишком много, и он начнет поступать непрерывно, — P2X7 уже не остановить. Через него начинает входить кальций и вытекать наружу калий, а это подает сигнал иммунным клеткам о том, что нужно мобилизовать тяжелую артиллерию и вовсю штамповать провоспалительные цитокины [34]. Делают они это при помощи создания больших молекулярных каруселей, которые называются инфламмасомами. Это вспомогательные белковые комплексы, которые участвуют в созревании и высвобождении интерлейкинов. Таким образом, рецептор P2X7 активирует сборку инфламмасомы NLRP3 и, следовательно, служит ключевым звеном в продукции интерлейкина-1β. Полюбоваться этим процессом в подробностях можно на видео:
Видео. Активация и сборка инфламмасомы NLRP3. Ее сборку и последующий синтез интерлейкина-1β напрямую связывают с активацией P2X7-рецепторов и оттоком через них ионов калия. Однако конкретный механизм запуска сборки остается пока под вопросом. На начальном этапе присутствующие в клетке субъединицы инфламмасомы активируют свои связывающие домены в ответ на сигнал стресса (в данном случае на повышенную активность P2X7-рецептора, вызванную всплеском уровня внеклеточного АТФ). Благодаря этому они становятся способны к олигомеризации и образованию сигнальных комплексов в виде колеса. LRR-домены (от англ. leucine-rich repeat), богатые остатками лейцина, располагаются снаружи колеса, в то время как белковые домены пирина (PYD — сокр. от англ. PYRIN domain) формируют круглую платформу в центре. Далее в дело вступает адаптерный белок ASC (сокр. от англ. Apoptosis-associated speck-like protein containing a CARD, где CARD — это caspase-recruitment domain). Этот белок имеет свой пириновый домен (PYD) со стороны N-конца, что позволяет ему соединяться с одноименным доменом инфламмасомы и формировать длинную ось на инфламмасомном кольце. На получившейся оси выступают наружу те самые C-концевые CARD-домены, необходимые для активации каспазы-1. Плавающие в цитозоле молекулы каспазы цепляются за эти домены и образуют активные димеры, в результате чего сами себя активирующие ферменты начинают бодро отрезать от незрелых про-интерлейкинов-1β куски белка, дающие им приставку про-. Каспаза-1 также отрезает куски от про-гасдермина-1, аналогичным образом переводя его в зрелую форму. Активированные молекулы гасдермина-1 формируют поры в клеточной мембране, что приводит к пироптозу — программируемой некротической гибели клетки в результате лизиса мембраны. Сформировавшиеся провоспалительные цитокины, в том числе полученные при помощи NLRP3-инфламмасомы интерлейкины-1β, вырываются наружу, сея хаос и неся воспаление окружающим тканям.
Как я уже говорил, при продолжительном действии большой концентрации агониста, в P2X7-рецепторе открывается большая пора, способная пропускать в клетку крупные растворимые молекулы размером до 900 дальтон. Длительное открытие такой макропоры, в зависимости от типа клеток, инициирует некроз или апоптоз. Причины запуска этих процессов пока непонятны, однако доказано, что открытие макропоры напрямую увеличивает продукцию провоспалительных цитокинов и процент гибнущих клеток. По некоторым версиям, макропора многократно усиливает отток ионов калия, который и так происходит при нормальной работе канала, либо же просто нарушает осмотическое давление клетки. Также продолжительная работа P2X7-рецептора ведет к слиянию мембран активированных макрофагов и образованию гигантских многоядерных клеток с их последующей гибелью. Такое свойство P2X7-рецептора может оказаться полезным, если в иммунные клетки закралась и спряталась болезнетворная бактерия (например, чумная палочка или сальмонелла). Ну или целый внутриклеточный паразит, такой, как риккетсия [41]. Механизм открытия большой поры пока остается загадкой и требует дальнейшего изучения. Ясно лишь, что это как-то связано с удлиненным C-концом и его сайтами связывания [42].

Рисунок 19. Воздействие высоких концентраций АТФ на P2X7-рецептор убивает опухолевые клетки у мышей. На рисунке изображен эксперимент по влиянию АТФ на подсаженные мышам опухоли гормонорезистентного рака простаты DU145. Исходя из свойств P2X7-рецептора, ожидалось, что регулярное воздействие большого количества АТФ станет цитотоксическим для клеток опухоли, либо активирует клеточный иммунитет у окружающих ее тканей. На фото продемонстрирован результат эксперимента спустя 60 дней — мышь снизу получала инъекции АТФ до 25 мМоль напрямую в опухоль. Мышь сверху не получала никакого лечения.
В желании использовать способности P2X7-рецептора в убийстве клеток высокие концентрации АТФ даже кололи лабораторным мышкам напрямую в подсаженные опухоли. В целом, помогало. Размер опухоли существенно уменьшался (рис. 19) [43].
Позже выяснилось, что раковые клетки умеют выкручиваться из любой ситуации и выставляют у себя на мембранах бракованные P2X7 (с точечными мутациями и аминокислотными заменами). Они спокойно пользуются благами ионного канала и кальциевого обмена, вот только мутировавший рецептор уже не может открыть смертоносную пору и уничтожить зловредную опухоль [44]. Да и нарушение работы рецептора в здоровых клетках может превратить их в опухолевые, если рецептор начнет нарушать гомеостаз и баланс ионов кальция [45]. Увы, в вопросах канцерогенеза всегда всё неоднозначно.
В общем, как у P2X7, так и у P2X4 в организме особые полномочия. Однако способность запускать воспаление и разрушать клетки — это весьма опасное молекулярное оружие, которое в любой момент может выйти из-под контроля. Разрушение клеток создаст выброс больших концентраций АТФ (внутри клетки его находится примерно в сто тысяч раз больше, чем снаружи). Эти большие концентрации опять запустят вышеупомянутые рецепторы, и те снова начнут вызывать выработку провоспалительных цитокинов и разрушать клетки. В том числе и здоровые. Это явление еще называют цитокиновым штормом. Страшное дело, натуральный порочный круг. Ученые до сих пор спорят: злодей или герой P2X7-рецептор в этой истории [46]. Ведь часто его удивительные свойства могут спасти организм, а не только навредить ему.
Нарушениям в работе этих рецепторов уже приписывают целый медицинский справочник заболеваний. В длинном списке есть всё, что касается боли, воспаления и гибели клеток: ревматоидный артрит, диабет, циррозы и фиброзы, аллергия, болезни Крона, Альцгеймера и Паркинсона, амиотрофический склероз, инфаркты и инсульты, рак... Даже психиатры стали засматриваться на P2X7. Повышенные уровни его синтеза в головном мозге наблюдаются у людей, страдающих депрессией, биполярным расстройством или острым психозом [47].
У невропатологов здесь тоже свой интерес. Выяснилось, что чрезмерная активация P2X4- и P2X7-рецепторов в микроглии (рис. 20) может привести к нейронному воспалению [48].
- Микроглия
- это резидентные макрофаги (то есть прикрепленные к «месту жительства» иммунные клетки-поглотители) в центральной нервной системе. Поскольку благодаря гематоэнцефалическому барьеру (ГЭБ) нервная система в норме стерильна, этим иммунным клеткам там особенно не разгуляться. Поэтому микроглия расширила свой функционал и выполняет для нейронов скорее роль техподдержки, регулируя их рост, но и о своей защитной функции при случае не забывает.

Рисунок 20. Микроглия. Клетки микроглии (окрашены зеленым) и нервные волокна (окрашены красным).
Это может обернуться гибелью нейронов и параличом. А еще стать причиной гипервозбудимости в рогах спинного мозга, что приводит к мучительным болевым синдромам. В том числе аллодинии и гипералгезии — неадекватной реакции на болевые и физические стимулы. Непросто жить, когда обычное поглаживание по руке взрывается в мозге болью тысячи кинжалов (рис. 21).

Рисунок 21а. Жизнь — боль. Эти картинки, на мой взгляд, лучше всего подходят для описания того, что испытывают люди со сверхактивацией P2X4- и P2X7-рецепторов в микроглии.

Рисунок 21б. Жизнь — боль. Эти картинки, на мой взгляд, лучше всего подходят для описания того, что испытывают люди со сверхактивацией P2X4- и P2X7-рецепторов в микроглии. На фото — Андраш Арато.
Конечно же, научные лаборатории активно ведут поиск блокаторов таких рецепторов, чтобы все эти неприятные заболевания предотвратить. Крупнейшие фармацевтические компании, такие как Abbot и AstraZeneca, от них не отстают [49]. Некоторые бизнес-проекты создаются исключительно для поиска и разработки лекарственных средств, действующих на P2X-рецепторы. Тут я имею в виду крупный исследовательский проект европейских ученых PurinesDX.
Блокаторы бывают двух видов. Первые конкурируют за место под солнцем с АТФ, по аналогии с кофеином для P1-рецепторов. Обманув P2X-рецептор, они занимают карманы для связывания АТФ, но, в отличие от привычного нуклеотида, не приводят к открытию канала. Карманы для АТФ выстланы положительно заряженными аминокислотными остатками лизина. Для этого хорошо подходят отрицательно заряженные молекулы, часто очень похожие на АТФ структурой. Так, например, блокатором P2X7-рецепторов служит периодатно окисленный АТФ (о-АТФ) (рис. 22а). Однако такие небольшие модификации природного агониста могут давать и обратный эффект: фенил-бензоат-АТФ (рис. 22б), напротив, открывает P2X7-рецептор в сорок раз эффективнее, чем «родной» эндогенный АТФ (то есть для открытия канала его потребуется в сорок раз меньше) [50].

Рисунок 22. Искусственно созданные модуляторы P2X7-рецепторов. а — Периодатно окисленный аденозинтрифосфат — эффективный блокатор P2X7-рецептора. б — Бензоильное производное АТФ — эффективный агонист P2X7-рецептора.
структуры созданы автором статьи при помощи ChemDraw Ultra 12
Второй тип блокаторов — это так называемые аллостерические лиганды (то есть соединения, которые находят свое место для связывания с рецептором и не конкурируют за него с АТФ) [62]. Часто такие вещества выполняют роль заглушек, что встают между субъединицами и мешают им раскрываться. В 2016 году тот же Каватэ получил кристаллическую структуру P2X7-рецептора в связке с различными синтетическими блокаторами [51]. Молекулы антагониста заняли полости, образованные ароматическими остатками между двумя субъединицами (рис. 23). Радует, что таких больших кармашков в этом месте нет у других типов рецепторов, а значит, мы можем подобрать очень избирательный блокатор именно для P2X7. Сюда прекрасно подойдут молекулы с плоскими ароматическими циклами, образуя сэндвич-подобную структуру, типичную для таких взаимодействий в белках.

Рисунок 23. Трехмерная модель P2X7-рецептора. В отдельных врезках в верхней части рисунка продемонстрированы виды сбоку и сверху на недавно найденный аллостерический сайт связывания, в который заходит один из современных антагонистов этого рецептора под шифром А804598. Этот аллостерический регулятор обладает двумя парами конденсированных ароматических гетероциклов, которые с любой стороны легко образуют сэндвич-подобные структуры с ароматическими аминокислотными остатками аллостерического сайта. Новые сайты связывания с антагонистом расположены совсем рядом с сайтами связывания АТФ и являются уникальным структурным явлением для данного типа рецепторов.
[51], рисунок адаптирован
Порой волшебная пилюля принимает неожиданные формы. Например, было показано, что эффективным блокатором P2X7-рецептора является BBG, он же бриллиантовый синий, он же кумасси (рис. 24б). Этот краситель хорошо известен всем, кто занимается молекулярной биологией. В XIX веке им красили шерсть, а сейчас он используется для окрашивания белков на электрофорезе. В далеком уже 2009 году в медицинском центре университета Рочестера, что в штате Нью-Йорк, проводилось одно исследование, в ходе которого несчастным мышкам повредили позвоночник, после чего их лечили инъекциями BBG [52]. Предполагалось, что вещество сумеет остановить вызванный травмой очаг воспаления и предотвратит гибель нервных тканей. Лечение оказалось весьма эффективным, некоторые мыши даже восстановили двигательные функции и смогли ходить, немного прихрамывая. Такой вот Иисус от мира молекулярной фармакологии. Контрольные мыши, которые не получали лечения, так и остались парализованными. Правда, у данного препарата был необычный побочный эффект — он активно окрашивал все кровеносные сосуды в насыщенный синий цвет (рис. 24а). Синими у мышей становились даже глаза. Фанатам «Дюны» должно понравиться.

Рисунок 24. BBG течет во мне, я един с BBG. а — Вполне ожидаемый побочный эффект, если решил лечить парализованных мышей известным красителем. б — Структура BBG.
История с BBG — это лишь один из сотен примеров. Ежегодно патентуется множество соединений, но большая их часть отсеивается на стадии клинических испытаний [49]. Одни оказываются токсичны, другие малоэффективны, третьи недостаточно селективны, а четвертые не могут проходить гематоэнцефалический барьер и достигать нервной системы. Хотя многие и показывают себя весьма эффективными in vitro и продаются коммерческими фирмами как селективные антагонисты для различных исследований. Тем не менее поиск идеальных блокаторов P2X-рецепторов всё еще остается актуальной задачей для современной фармакологии.
P2Y и P1: пурины на службе у GPCR
Скажем еще пару слов о системе, разработанной в ходе эволюции для продуктов распада АТФ. Они представлены в виде GPCR — метаботропных рецепторов, родственных светочувствительному белку родопсину [53]. Иначе говоря, это такие рецепторы, которые, активируясь, воздействуют на универсальные внутриклеточные посредники — G-белки. Таким образом они запускают выделение небольших, но очень активных молекул. Эти молекулы — так называемые вторичные мессенджеры — заметно влияют на клеточные процессы и инициируют в клетках каскады биохимических реакций.
Первыми на очереди стоят P2Y-рецепторы. Выглядят они вполне типично для GPCR: семь спиралей, змейкой пронизывающих насквозь клеточную мембрану (рис. 25). При активации они изменяют свою конформацию и таким образом переключают G-белки. Всего их в пределах семейства Y восемь разновидностей (с непростой нумерацией): Y1, Y2, Y4, Y6, Y11, Y12, Y13, Y14. Словно корпорация Майкрософт, нумерующая свои операционные системы. Это объясняется историей открытия и тем, что при их клонировании некоторые GPCR-рецепторы ошибочно заносились в пуриновые. Позже классификация устаканилась, и лишнее пришлось убрать.

Рисунок 25. P2Y12-рецептор со своим агонистом АДФ (фиолетовый). Типичный GPCR: серпантин из семи соединенных спиралей, пронизывающий клеточную мембрану (серая полоса). N-конец и три петли снаружи клетки, С-конец и три петли внутри.
«Игреки» распространены практически во всех типах тканей и выполняют ряд важных функций [54]. В нервной системе они регулируют нейрогенез, рост и дифференцировку нервных клеток. В поджелудочной железе одни P2Y стимулируют выработку инсулина, а другие, наоборот, ее прекращают [55]. В кровеносной системе запускают агрегацию тромбоцитов и повышают свертываемость крови. Кстати, именно Бёрнстоку удалось открыть и помочь вывести на рынок препарат с благозвучным названием клопидогрел, который как раз блокирует такие рецепторы (а именно P2Y12) и предотвращает образование тромбов [56]. Некоторые P2Y-рецепторы способствуют расслаблению гладкой мускулатуры в стенках кровеносных сосудов (включая сосуды головного мозга), тем самым регулируя наше с вами кровяное давление. Чем не мишень для лечения мигрени и различных сердечно-сосудистых заболеваний [57]?
А еще P2Y (вместе с P2X) вовлечены в процессы воспаления и иммунного ответа [58]. А где иммунный ответ, там, вероятно, и злоупотребление его полномочиями, которое приводит к разрушению «попавших под раздачу» клеток. Разрушенные клетки со временем будут замещены соединительной тканью [59]. Врачи называют этот процесс фиброзом. Пользуясь случаем, передаем горячий привет циррозу печени [60] и синдрому сухого глаза [61] (крайне неприятное заболевание, которое рано или поздно может привести своих обладателей к слепоте). Как вы уже поняли, такая широкая распространенность P2Y-рецепторов и многообразие их свойств — это просто кладезь для разработки лекарственных препаратов. Что, впрочем, справедливо для большинства GPCR — читайте об этом в статье: «Аллостерические регуляторы GPCR: ключи от всех замков» [62].

Рисунок 26. Кофеин неспроста напоминает молекулу аденозина без рибозы. Этого двойника растения шлифовали миллионы лет, чтобы изводить докучающих насекомых, блокируя их аденозиновые рецепторы. А уже потом мы стали делать эспрессо.
Другими важными GPCR в пуриновой системе сигнализации служит семейство P1-рецепторов. Оно состоит из четырех представителей — P1, P2A, P2B и P3. Эти рецепторы отвечают за самые разнообразные процессы. В нервной системе их работа лежит в сфере пресинаптического торможения. Они также вызывают у нас ощущение усталости, когда аденозина в ЦНС накопится слишком много в результате нервной деятельности и распада АТФ. Благодаря P1-рецепторам мозг понимает, что ему пора отдохнуть и обновить свои системы при помощи сна. Ну а подавляются эти рецепторы, как уже было сказано ранее, метилксантинами. Ведь метилксантины структурно очень похожи на аденозин без рибозы. Настолько похожи, что рецепторы не видят подмены и заглатывают фальшивку, а на деле заглушку. К метилксантинам относится и хорошо знакомый нам кофеин (рис. 26). Именно так кофе помогает нам взбодриться. Если сильно пристраститься к кофеину, нервная система начинает производить больше аденозиновых рецепторов. Однако если прекратить употреблять этот кофейный алкалоид, то аденозин займет освободившиеся места на этой куче рецепторов, и мы будем острее ощущать сонливость без привычной дозы бодрящего напитка. Более подробно о действии кофе читайте в статье «Влияние кофеина» на «Биомолекуле» [63].
Также интересный вывод сделали ученые при изучении акупунктурных методик лечения (рис. 27). Утверждается, что иглоукалывание стимулирует локальный выброс аденозина, который воздействует на P1А1-рецептор и тем самым помогает снять острую боль и воспаление. Конечно, и тут не обошлось без АТФ. Сомневались в наличии научных доказательств в пользу иглоукалываний? И правильно, в общем-то, делали. Но, кажется, теперь такие доказательства имеют место [64].

Рисунок 27. Процедура иглоукалывания. По одному из предложенных учеными механизмов — игла стимулирует локальный выход АТФ, который разлагается внеклеточными ферментами до аденозина. Аденозин воздействует на пресинаптические P1A1-рецепторы, которые ингибируют работу (то есть препятствуют потенциалу действия) периферических нервных волокон, отвечающих за передачу болевого сигнала в центральную нервную систему.
Помимо этого, у аденозиновых P1-рецепторов в организме множество функций, однако мы не будем останавливаться на них. Подробнее об этих рецепторах и о том, как их обманывают растительные метаболиты, на «Биомолекуле» уже есть интересная статья, с которой я всем советую ознакомиться: «Аденозиновые рецепторы: история великого обмана» [65].
У эволюционных истоков
Пуриновые и пиримидиновые нуклеотиды возникли на Земле еще в пребиотическую эпоху. И они же стали причиной эпохи биотической [66], [67]. Ведь без нуклеотидов не появились бы ДНК и РНК, которые составляют основу жизни в привычном нам понимании. АТФ уже на ранних этапах стал главным источником энергии для всех живых организмов. Это был невероятно важный эволюционный выбор. Благодаря нему сформировались целые ферментативные системы для использования АТФ в качестве главной рабочей лошадки, что, в свою очередь, повлекло за собой создание универсальной сигнальной системы, основанной на ионах кальция (Ca2+).
Почему опять кальций, спросите вы? Из-за чего он стал таким важным? Дело в том, что, по выбранному природой сценарию, этого кальция в цитоплазме должно находиться крайне мало. Ведь при большом количестве могут образоваться нерастворимые фосфаты кальция, а использовать по уши фосфатированный АТФ для энергетики клетки стало бы затруднительно. Соответственно, поступающий кальций должен был задействоваться в физиологических процессах и утилизироваться из клетки. Это и повлияло на его роль в качестве вторичного мессенджера. Сюда также вписывается теория о повышенной щелочной среде первичного океана (а жизнь на Земле начинала развиваться именно в таких условиях), в котором ионов кальция было мало (дело, опять же, в плохой растворимости гидроксидов кальция). В наших клетках большая доля кальция сосредоточена в кальциевых депо эндоплазматического ретикулума, из которых он выделяется для передачи сигнала, а также он может поступать снаружи благодаря ионным каналам вроде P2X.
Примитивным организмам уже на первых порах понадобились рецепторы для оценки окружающей среды, будь то древний океан или вулканические гейзеры (для теплолюбивых архей). По мере появления многоклеточности также встал вопрос о передаче информации от одной клетки к другой. Тут мы можем строить лишь догадки, но накопленные знания по биологии и филогении примитивных организмов позволяют нам сделать кое-какие выводы. По-крайней мере, можно сойтись во мнении, что первыми передатчиками стали ионотропные рецепторы, а методом межклеточной коммуникации — обычная диффузия.
Собственно, нейромедиаторы и сейчас гуляют через синаптическую щель благодаря диффузии [68]. На роль диффундирующих передатчиков выбор был невелик — ионы и мелкие растворимые молекулы, присутствующие в самих клетках. Чего-чего, а ионов первобытный океан содержал в избытке, поэтому их (кроме, вероятно, протонов) можно исключить. В качестве межклеточных посыльных годились растворимые аминокислоты и их производные (одними из самых древних считаются рецепторы глутамата); протоны, которые клетки накапливали в процессе метаболизма; а также некоторые газообразные молекулы, такие как монооксид азота (NO). Ну и, конечно же, пурины, особенно пурины фосфатированные, благодаря их хорошей растворимости. Есть предположение, что первые АТФ-рецепторы использовались только для реакции на сигналы стресса при разрушении клеток, а литический путь попадания этого нуклеотида во внеклеточную среду был единственно возможным. Однако по мере усложнения организмов эта система развилась до выполнения и других функций, а АТФ стал выделяться направленно и дозированно.
На протяжении всей статьи мы говорили в основном о рецепторах позвоночных. Однако если копнуть немного глубже, пуриновые рецепторы найдутся даже у бактерий. Грибы и растения, моллюски и насекомые, анемоны и кольчатые черви — все они имеют свои пуринорецепторы, выполняющие самые разнообразные функции [69].
У ракообразных, например, пуринергические рецепторы играют важнейшую роль в обонянии и вкусе. Омары (рис. 28) могут учуять в морской воде даже мельчайшие крохи АТФ и АМФ. Вкусно пахнущий аденозинтрифосфат сигнализирует им о раненой добыче, а вот его деградировавшая версия в виде монофосфата будет отталкивать разборчивых членистоногих, сообщая о том, что предполагаемая добыча уже испортилась.

Рисунок 28. Европейский омар (Homarus gammarus). Это ракообразное обладает острым нюхом на пурины. Запах АТФ будет для омаров всё равно что для нас запах выпечки или жареного мяса и будет сообщать им о свежей добыче.
Многие планктонные организмы обзавелись целыми наборами дефосфорилирующих ферментов, чтобы быстро разлагать выделяемые нуклеотиды и маскироваться от хищников. АТФ также важен для восприятия пищи и формирования пищевого поведения у различных членистоногих паразитов, таких как муха цеце, комары или клещи. Активация АТФ-рецепторов в эритроцитах отвечает у них за чувство насыщения и говорит, что крови эти паразиты выпили достаточно.
А вот АТФ-рецепторы растений были клонированы и охарактеризованы сравнительно недавно — в 2014 году [70]. Назвали их DORN-1 и DORN-2. Они выполняют в растениях функции стресса, схожие с аналогичными у P2X-рецепторов животных. Реагируя на повышенные концентрации внеклеточного АТФ, в клетки растений входит кальций, который запускает митоген-активированную протеинкиназу и приводит к повышенной продукции активных форм кислорода. Всё это — ярчайшие маркеры клеточного стресса, поэтому растение начинает активно экспрессировать защитные гены. В общем, теория о первоначальной роли АТФ-рецепторов в очередной раз подтверждается.
Также ученые доказали, что хищное растение венерина мухоловка захлопывает свою пасть, реагируя на молекулы АТФ [71]. Попав в ловушку, насекомое задевает механочувствительные волоски, которые стимулируют выброс АТФ во внеклеточную среду. Под действием этого нейромедиатора срабатывает потенциал действия, благодаря которому растение закрывает створки, чтобы превратить муху в питательное удобрение (рис. 29).

Рисунок 29. Венерина мухоловка ловит муху
Единообразие функций и строения пуринорецепторов среди живых организмов поражает. Те же P2X-рецепторы можно найти и у одноклеточных эукариот, хоть и сходство с таковыми у млекопитающих будет довольно низким. Тем не менее такие рецепторы всё еще сохраняют свою микромолярную чувствительность к АТФ, а появились они более миллиарда лет назад. Считается, что именно ионные каналы по типу P2X-рецепторов стали первыми ласточками пуринергической передачи сигнала, а уже значительно позже за ними последовали метаботропные рецепторы, сопряженные с G-белком, такие как P0, P1 или P2Y. Изучая и сравнивая гены, кодирующие пуринорецепторы в различных организмах, ученые смогут многое почерпнуть о времени эволюционного расхождения различных форм жизни на планете.
Пуринергическая сигнализация, опосредованная АТФ и родственными нуклеотидами, присутствует на всех этапах эволюции живых существ и стоит буквально у истоков жизни.
Благодарности
Автор благодарит своего научного руководителя, д.б.н. Аминина Дмитрия Львовича за наставление на путь исследований в данной тематике и неоценимую помощь в работе. Эта статья посвящается памяти Джеффри Бёрнстока, который ушел из жизни 2 июня 2020 года, прожив долгую и плодотворную научную жизнь.
Литература
- Внимание! Разыскивается предок митохондрий!;
- Geoffrey Burnstock. (2014). Purinergic signalling: from discovery to current developments. Experimental Physiology. 99, 16-34;
- 12 методов в картинках: нейробиология;
- Тетродотоксин — история элегантного убийцы;
- G. Burnstock, G. Campbell, M. J. Rand. (1966). The inhibitory innervation of the taenia of the guinea-pig caecum. The Journal of Physiology. 182, 504-526;
- A. N. Drury, A. Szent-Györgyi. (1929). The physiological activity of adenine compounds with especial reference to their action upon the mammalian heart1. The Journal of Physiology. 68, 213-237;
- Pamela Holton. (1959). The liberation of adenosine triphosphate on antidromic stimulation of sensory nerves. The Journal of Physiology. 145, 494-504;
- G. BURNSTOCK, G. CAMPBELL, D. SATCHELL, ANNE SMYTHE. (1970). Evidence that adenosine triphosphate or a related nucleotide is the transmitter substance released by non-adrenergic inhibitory nerves in the gut. British Journal of Pharmacology. 40, 668-688;
- Burnstock G. (1972). Purinergic nerves. Pharmacological reviews. 3, 509–581;
- Burnstock G. A basis for distinguishing two types of purinergic receptor. In: Cell Membrane Receptors for Drugs and Hormones: A Multidisciplinary Approach // ed. by Straub R.W. and Bolis L. NY: Raven Press, 1978. P. 107–118;
- Великому комбинатору и не снилось: комбинаторика токсинов пауков;
- Яды — высокоточное оружие: компьютерное исследование природных нейротоксинов;
- G. Burnstock, C. Kennedy. (1985). Is there a basis for distinguishing two types of P2-purinoceptor?. General Pharmacology: The Vascular System. 16, 433-440;
- Молекулярное клонирование, или как засунуть в клетку чужеродный генетический материал;
- 12 методов в картинках: генная инженерия. Часть II: инструменты и техники;
- Dominik Thimm, Melanie Knospe, Aliaa Abdelrahman, Miguel Moutinho, Bernt B. A. Alsdorf, et. al.. (2013). Characterization of new G protein-coupled adenine receptors in mouse and hamster. Purinergic Signalling. 9, 415-426;
- Constanze Kaebisch, Dorothee Schipper, Patrick Babczyk, Edda Tobiasch. (2015). The role of purinergic receptors in stem cell differentiation. Computational and Structural Biotechnology Journal. 13, 75-84;
- Toshimitsu Kawate, Jennifer Carlisle Michel, William T. Birdsong, Eric Gouaux. (2009). Crystal structure of the ATP-gated P2X4 ion channel in the closed state. Nature. 460, 592-598;
- L. E. Browne, J. P. M. Nunes, J. A. Sim, V. Chudasama, L. Bragg, et. al.. (2014). Optical control of trimeric P2X receptors and acid-sensing ion channels. Proceedings of the National Academy of Sciences. 111, 521-526;
- Emily Caseley, Stephen Muench, Sebastien Roger, Hong-Ju Mao, Stephen Baldwin, Lin-Hua Jiang. (2014). Non-Synonymous Single Nucleotide Polymorphisms in the P2X Receptor Genes: Association with Diseases, Impact on Receptor Functions and Potential Use as Diagnosis Biomarkers. IJMS. 15, 13344-13371;
- Luiz Alves, João da Silva, Dinarte Ferreira, Antonio Fidalgo-Neto, Pedro Teixeira, et. al.. (2014). Structural and Molecular Modeling Features of P2X Receptors. IJMS. 15, 4531-4549;
- Guy Vassort. (2001). Adenosine 5′-Triphosphate: a P2-Purinergic Agonist in the Myocardium. Physiological Reviews. 81, 767-806;
- Наконец-то: противозачаточная таблетка для мужчин!;
- C. W. White, Y.-T. Choong, J. L. Short, B. Exintaris, D. T. Malone, et. al.. (2013). Male contraception via simultaneous knockout of 1A-adrenoceptors and P2X1-purinoceptors in mice. Proceedings of the National Academy of Sciences. 110, 20825-20830;
- Geoffrey Burnstock. (2017). Purinergic Signaling in the Cardiovascular System. Circ Res. 120, 207-228;
- Sylvain Hugel, Rémy Schlichter. (2000). Presynaptic P2X Receptors Facilitate Inhibitory GABAergic Transmission between Cultured Rat Spinal Cord Dorsal Horn Neurons. J. Neurosci.. 20, 2121-2130;
- Larry Rodriguez, Catherine Yi, Cameron Chu, Quentin Duriez, Sharyse Watanabe, et. al.. (2020). Cross-Talk between P2X and NMDA Receptors. IJMS. 21, 7187;
- D. Yan, Y. Zhu, T. Walsh, D. Xie, H. Yuan, et. al.. (2013). Mutation of the ATP-gated P2X2 receptor leads to progressive hearing loss and increased susceptibility to noise. Proceedings of the National Academy of Sciences. 110, 2228-2233;
- Brederson J.D. and Jarvis M.F. (2008). Homomeric and heteromeric P2X3 receptors in peripheral sensory neurons. Current opinion in investigational drugs. 9(7), 716–725;
- Oleg O. Krishtal. (2009). Novel peptide from spider venom inhibits P2X3 receptors and inflammatory pain. Ann Neurol.. NA-NA;
- R. Sathanoori, F. Rosi, B. J. Gu, J. S. Wiley, C. E. Müller, et. al.. (2015). Shear stress modulates endothelial KLF2 through activation of P2X4. Purinergic Signalling. 11, 139-153;
- Ronghua Yang, Bruce T. Liang. (2012). Cardiac P2X 4 Receptors. Circ Res. 111, 397-401;
- Luca Antonioli, Corrado Blandizzi, Matteo Fornai, Pál Pacher, H. Thomas Lee, György Haskó. (2019). P2X4 receptors, immunity, and sepsis. Current Opinion in Pharmacology. 47, 65-74;
- Francesco Di Virgilio, Diego Dal Ben, Alba Clara Sarti, Anna Lisa Giuliani, Simonetta Falzoni. (2017). The P2X7 Receptor in Infection and Inflammation. Immunity. 47, 15-31;
- Airi Rump, Olli Pekka Smolander, Sirje Rüütel Boudinot, Jean M. Kanellopoulos, Pierre Boudinot. (2020). Evolutionary Origin of the P2X7 C-ter Region: Capture of an Ancient Ballast Domain by a P2X4-Like Gene in Ancient Jawed Vertebrates. Front. Immunol.. 11;
- Markus Schneider, Kirsten Prudic, Anja Pippel, Manuela Klapperstück, Ursula Braam, et. al.. (2017). Interaction of Purinergic P2X4 and P2X7 Receptor Subunits. Front. Pharmacol.. 8;
- J L Gordon. (1986). Extracellular ATP: effects, sources and fate. Biochemical Journal. 233, 309-319;
- A. Surprenant, F. Rassendren, E. Kawashima, R. A. North, G. Buell. (1996). The Cytolytic P2Z Receptor for Extracellular ATP Identified as a P2X Receptor (P2X7). Science. 272, 735-738;
- Robin Kopp, Anna Krautloher, Antonio Ramírez-Fernández, Annette Nicke. (2019). P2X7 Interactions and Signaling – Making Head or Tail of It. Front. Mol. Neurosci.. 12;
- Alison Gartland, Kristen K Skarratt, Lynne J Hocking, Claire Parsons, Leanne Stokes, et. al.. (2012). Polymorphisms in the P2X7 receptor gene are associated with low lumbar spine bone mineral density and accelerated bone loss in post-menopausal women. Eur J Hum Genet. 20, 559-564;
- Luiz Eduardo Baggio Savio, Robson Coutinho-Silva. (2019). Immunomodulatory effects of P2X7 receptor in intracellular parasite infections. Current Opinion in Pharmacology. 47, 53-58;
- Francesco Di Virgilio, Günther Schmalzing, Fritz Markwardt. (2018). The Elusive P2X7 Macropore. Trends in Cell Biology. 28, 392-404;
- Geoffrey Burnstock, Francesco Di Virgilio. (2013). Purinergic signalling and cancer. Purinergic Signalling. 9, 491-540;
- Julian A. Barden. (2014). Non-Functional P2X7: A Novel and Ubiquitous Target in Human Cancer. J Clin Cell Immunol. 05;
- Romain Lara, Elena Adinolfi, Catherine A. Harwood, Mike Philpott, Julian A. Barden, et. al.. (2020). P2X7 in Cancer: From Molecular Mechanisms to Therapeutics. Front. Pharmacol.. 11;
- Luiz E. B. Savio, Paola de Andrade Mello, Cleide Gonçalves da Silva, Robson Coutinho-Silva. (2018). The P2X7 Receptor in Inflammatory Diseases: Angel or Demon?. Front. Pharmacol.. 9;
- Anindya Bhattacharya. (2018). Recent Advances in CNS P2X7 Physiology and Pharmacology: Focus on Neuropsychiatric Disorders. Front. Pharmacol.. 9;
- Tsuda Makoto, Tozaki-Saitoh Hidetoshi, Inoue Kazuhide. (2012). P2X4R and P2X7R in neuropathic pain. WIREs Membr Transp Signal. 1, 513-521;
- Jin-Hee Park, Yong-Chul Kim. (2017). P2X7 receptor antagonists: a patent review (2010–2015). Expert Opinion on Therapeutic Patents. 27, 257-267;
- Mark T. Young, Pablo Pelegrin, Annmarie Surprenant. (2007). Amino Acid Residues in the P2X7 Receptor that Mediate Differential Sensitivity to ATP and BzATP. Mol Pharmacol. 71, 92-100;
- Akira Karasawa, Toshimitsu Kawate. (2016). Structural basis for subtype-specific inhibition of the P2X7 receptor. eLife. 5;
- W. Peng, M. L. Cotrina, X. Han, H. Yu, L. Bekar, et. al.. (2009). Systemic administration of an antagonist of the ATP-sensitive receptor P2X7 improves recovery after spinal cord injury. Proceedings of the National Academy of Sciences. 106, 12489-12493;
- Зрительный родопсин — рецептор, реагирующий на свет;
- Kenneth A. Jacobson, Esmerilda G. Delicado, Christian Gachet, Charles Kennedy, Ivar Kügelgen, et. al.. (2020). Update of P2Y receptor pharmacology: IUPHAR Review 27. Br J Pharmacol. 177, 2413-2433;
- A. Farret, M. Vignaud, S. Dietz, J. Vignon, P. Petit, R. Gross. (2004). P2Y Purinergic Potentiation of Glucose-Induced Insulin Secretion and Pancreatic Beta-Cell Metabolism. Diabetes. 53, S63-S66;
- Marco Cattaneo. (2011). The platelet P2Y12 receptor for adenosine diphosphate: congenital and drug-induced defects. Blood. 117, 2102-2112;
- Akiyuki Nishimura, Caroline Sunggip, Sayaka Oda, Takuro Numaga-Tomita, Makoto Tsuda, Motohiro Nishida. (2017). Purinergic P2Y receptors: Molecular diversity and implications for treatment of cardiovascular diseases. Pharmacology & Therapeutics. 180, 113-128;
- Diana Le Duc, Angela Schulz, Vera Lede, Annelie Schulze, Doreen Thor, et. al.. (2017). P2Y Receptors in Immune Response and Inflammation. Advances in Immunology. 85-121;
- Tobias Müller, Susanne Fay, Rodolfo Paula Vieira, Harry Karmouty-Quintana, Sanja Cicko, et. al.. (2017). The purinergic receptor subtype P2Y2 mediates chemotaxis of neutrophils and fibroblasts in fibrotic lung disease. Oncotarget. 8, 35962-35972;
- Geoffrey Burnstock, Byron Vaughn, Simon C. Robson. (2014). Purinergic signalling in the liver in health and disease. Purinergic Signalling. 10, 51-70;
- Simon Skalicky, Oliver Lau, Chameen Samarawickrama. (2014). P2Y2 receptor agonists for the treatment of dry eye disease: a review. OPTH. 327;
- Аллостерические регуляторы GPCR: ключи от всех замков;
- Влияние кофеина;
- Jin-Rong He, Shu-Guang Yu, Yong Tang, Peter Illes. (2020). Purinergic signaling as a basis of acupuncture-induced analgesia. Purinergic Signalling. 16, 297-304;
- Аденозиновые рецепторы: история великого обмана;
- CYRIL PONNAMPERUMA, CARL SAGAN, RUTH MARINER. (1963). Synthesis of Adenosine Triphosphate Under Possible Primitive Earth Conditions. Nature. 199, 222-226;
- Erik Galimov. (2009). Concept of Sustained Ordering and an ATP-related Mechanism of Life’s Origin. IJMS. 10, 2019-2030;
- Как происходит выделение нейромедиатора;
- Alexei Verkhratsky, Geoffrey Burnstock. (2014). Biology of purinergic signalling: Its ancient evolutionary roots, its omnipresence and its multiple functional significance. BioEssays. 36, 697-705;
- J. Choi, K. Tanaka, Y. Cao, Y. Qi, J. Qiu, et. al.. (2014). Identification of a Plant Receptor for Extracellular ATP. Science. 343, 290-294;
- Kiwamu Tanaka, Simon Gilroy, Alan M. Jones, Gary Stacey. (2010). Extracellular ATP signaling in plants. Trends in Cell Biology. 20, 601-608;
- Формирование мембранного потенциала покоя.



Комментарии
0Чтобы оставить комментарий, необходимо
войти